

브라질의 커피 잎 녹병(CLR)에 대한 본 리뷰에서는 우리는 다음과 같은 내용들을 보고한다.
(i) 브라질에서 CLR의 역사적 도입과 초기의 통제 조치;
(ii) 이 질병에 유리한 환경 조건 및 연중 시기;
(iii) CLR 저항성 재배품종 개발에 사용되는 육종 방법 및 전략;
(iv) CLR 저항성의 수준, 원인 및 유형;
(v) 브라질 저항성 재배품종의 개발; 그리고
(vi) 화학적 및 재배적 통제 방법.
대부분의 농장은 Catuaí 및 Mundo Novo 그룹과 같은 취약성 재배품종들로 재배된다.
브라질 연구 기관들에서는 다양한 수준의 저항성을 지닌 수십종의 재배품종들을 개발했으며, 새로운 저항성 재배품종의 재배를 크게 늘렸다.
CLR 저항성의 주요 원인은 Híbrido de Timor, Icatu, SH3 유전자를 보유하는 BA 시리즈 및 에티오피아 야생 커피들의 유전자형들이다. Sarchimor와 SH3-보유 유전자형들에서는 높은 CLR 저항성이 여전히 관찰된다. 중간 정도의 CLR 저항성은 에티오피아 야생 커피와 Sarchimor 및 Icatu 파생물에서 관찰되며, 정성적 저항성은 Hemileia vastatrix 균종들에 의해 대체되었다(밀렸다).
접촉성 살균제, 중간 살균제, 전신 살균제가 브라질에서 화학적 방제에 사용된다.
브라질의 CLR 발병은 11월 우기가 시작된 후 증가하기 시작하여 6월에 최고조에 도달하고 8월까지 높은 수준을 유지한다. 따라서 화학적 방제는 일반적으로 12월부터 4월까지 적용된다.

아라비카 커피는 브라질에 1727년에 도입되었고, 곧 이 나라의 역사와 경제적 발전에 직결되는 중요한 농산물이 되었다. 그러나, 이 농업은 1970년 1월 브라질 연구자 Arnaldo Gomes Medeiros에 의해, 브라질 Bahiha 남부에서 식별된 최초의 커피 잎 녹병 관찰에 의해 흔들렸다 [1]. CLR의 병인체는 생물영양 기생 곰팡이균(biotrophic fungus)인 Hemileia vastatrix이며, 이는 Berkeley and Broome이 1869년에 기술했다 [2,3].
이 질병의 기원을 설명하는 두 개의 가설이 있다. 첫 번째는 아프리카에서 가져온 코코아 묘목에서 나왔다는 것이다. 두 번째는 그것이 역시 아프리카에서 대서양을 가로질러 높은 고도의 기류에 의해 운반된 포자를 통해 도착했다고 하는 것이다 [4]. 그때까지 브라질 기술자들은 문헌 참조를 통해서만 이 질병에 대해 알고 있었다. 따라서 CLR이 국내 커피 작물을 파괴할 것이라는 상당한 우려가 있었고, 이로 인해 브라질 정부는 브라질 커피 연구소와 농업부를 통해 문제를 평가하기 위해 해외 전문가 방문을 장려했다; 특히 포르투갈 오에이라스에 위치한 Coffee Rusts Research Center (CIFC, Centro de Investigação das Ferrugens do Cafeeiro)의 과학자 Branquinho d'Oliveira의 방문을 추진했다.
문제의 심각성을 고려하여 브라질 정부는 곧 일련의 방제 대책을 수립하고 커피 녹병 방제 프로그램을 시행했다. 첫 번째 조치는 질병 발생 근절을 위해 피해 지역을 진단하는 것이었다. 조사는 Bahiha에서 시작했는데, 감염 지역을 더 남쪽에 위치한 주요 생산 지역과 분리하기 위해 약 50km 너비의 안전구역을 구축함으로써 시작되었다. 감염된 지역에서 발견된 모든 커피 나무들은 근절되었다. 동시에, 감염된 나무를 자르고 태워서 초기에 근절되었던 새로운 질병 발생을 식별하기 위해 CLR에 대한 정보 캠페인이 수행되었다. 이후 경우에 따라 고엽제(Paraquat)나 제균제 (Pyracarbolid)를 적용했다 [4].
CLR 병소를 근절 및 격리하고 확산을 방지하려는 노력에도 불구하고 질병은 전국적으로 빠르게 퍼졌다. 짧은 시간 안에 CLR은 1970년 6월 미나스 제라이스 남부, 1971년 1월 상파울루 등 가장 중요한 브라질 커피 지역에서 보고되었다. 그 당시 비행기에 장착된 트랩은 지상 최대 1,000m 높이에서 H. vastatrix 포자의 존재를 감지했으며, 바람의 작용이 CLR의 장거리 확산 요인임을 드러냈다.
녹 발생을 근절하고 격리하는 정책이 실현 불가능하다는 것을 확인한 후, 연구, 기술 지원 및 특정 재정 지원을 기반으로 CLR과의 공존 프로그램이 시작되었다. 연구 기관은 두 가지 주요 통제 전략에 중점을 두었다. 첫 번째는 매력적인 단기 전략인 화학적 방제이다. 대부분의 브라질 커피 작물은 상대적으로 높은 고도에 재배되며 건기가 뚜렷하여 아프리카, 아시아 및 태평양 지역의 커피 작물보다 화학적 방제에 더 적합하다. 두 번째 주요 통제 전략은 유전적 육종 (genetic breeding)이다.
초기 방제 시도 직후, Monaco [5]는 효율적인 화학적 방제 방법이 가능하기 때문에 녹 방제가 기술적인 문제보다 경제적인 문제라고 가정했다. 따라서 브라질에서는 질병 통제와 커피 생산 간의 절충안이 확립되었으며, 이로 인해 수확량이 적은 작물이 제거되고 커피 작물에 통합된 해충 및 질병 통제 관리 루틴이 시작되었다. 첫째, 살균제 수입이 증가했고, 브라질 커피 재배환경에 적합한 살포 장비가 개발됐다. 권장 사항은 헥타르 당 5kg의 옥시염화구리 (copper oxychloride)를 4번 뿌리는 것이었고, 당시 전체 커피 작물을 보호하려면 약 50,000톤의 살균제가 필요했다. 이로 인해 자본 투자 및 인건비 등으로 2억 달러 이상의 추가 비용이 발생했다.
이에 따라 지역 연구 센터가 설치되고, 기술 지원 네트워크, 인프라 구축 및 인력 교육이 확장되고, 커피재배 농업기후 구역(coffee-growing agroclimatic zoning)이 설정되었으며, CLR 제어를 위한 새로운 관리 방식을 갖춘 기술 및 경제 연구가 발달되었다. 1969년에 새로운 커피 농장을 장려하기 위해 “Coffee Crop Renewal and Reinvigorating Plan”이 만들어졌으며 1980년까지 운영되었다. 거의 20억 그루에 달하는 커피나무에 대한 리노베이션에 자금이 조달되고 기술적으로 지원되었다. 연구에 의해 개발 조정된 주요한 기술적 향상들이 통합되어진 새로운 크롭들이, 새로운 플랜팅을 위한 구역 지정, 새로운 식재 간격, 새로운 재배 품종들, 더욱 합리적이고 기술적인 관리, 그리고 통합관리 프로그램 내에서 체계적인 커피 병충해 통제의 도입을 강조하면서 지역적 기술지원을 통해 생산자들에게 제공되었다. 그 목표는 생산성을 높이고 생산 비용을 줄이는 것이었다.
CLR은 브라질의 커피 재배에 영향을 미치는 주요 질병이지만, 이 질병의 확산은 브라질 커피 작물에 긍정적인 영향을 미쳤다. 왜냐하면 CLR이 새로운 기술의 생성 및 채택을 통해 현대화를 촉진하고 보다 효율적인 관리 관행을 촉진하여 커피 경쟁력이 향상되었고 새로운 생산 지역으로의 생산 및 확장이 이루어졌기 때문이다. 1980년대 초 브라질에는 34억 개체로 구성된 커피 나무 스탠드가 있었는데, 이는 1970년보다 55% 증가한 수치이며, 평균 수확량은 60kg 그린 커피 2,500만 bags으로, 이는 1970년 수확 bags에 비해 연간 약 500만 bags이 증가한 것이다 [6]. 이러한 생산량 증가는 1975년 심한 서리로 인한 심각한 피해에도 불구하고 이루어졌으며, 이로 인해 1976년 커피 수확량이 단 600만 bags으로 감소했다. 이후 수십 년 동안 브라질의 커피 재배 농장은 새로운 기술로 인해 생산성을 계속해서 확장하고 증가시켰다. 현재 브라질은 약 220만 헥타르의 면적에서 60kg 생두 bag을 연간 약 4,500만~6,500만 bags 생산하며, 평균 생산량은 아라비카가 헥타르 당 32.18 bags, 로부스타가 헥타르당 38.78 bags이다 [7]. 브라질은 경쟁력 있는 가격과 대량생산 능력으로 국제적으로 인정받고 있지만, 최근에는 스페셜티 커피의 명망 높은 산지로도 두각을 나타내고 있다. 브라질은 또한 세계에서 두 번째로 큰 음료 소비국으로, 2021년 예상 소비량은 2,353만 bags (세계 소비의 14.4%)로 미국(16%)을 능가한다 [8,9].
브라질에서, CLR은 재배품종의 저항성 수준, 이 질병에 유리한 기후조건들, 그리고 관리 조치들에 따라 커피 생산에서 50%까지의 손실을 유발할 수 있으며, 멕시코, 코스타리카, 엘살바도르와 같은 다른 커피 생산국가들에서도, CLR에 의한 커피 소출 손실은 매우 중요하다 [10-13].

Hemileia 屬은 Basidiomycota 門, Pucciniomycetes 綱, 그리고 Pucciniales 目의 구성원이다. (擔子菌 門, 綠菌 綱, 綠菌 目) Hemileia라는 이름은 분산과 감염을 돕는 urediniospores (하포자)의 반쯤 매끄러운(반 거친) 특징적인 형태를 반영한다.
H. Vastatrix urediniospores (하포자)는 reniform (腎腸形) (28~36 × 18~28μm)이며, 볼록한 면에는 무사마귀가 있고(warted), 곧은 또는 오목한 면에서는 매끄러운, 琉璃質 壁(hyaline wall)으로 덮여 있으며 두께는 1μm이다 [3,14,15] .
첫 번째 CLR 증상은 작고 연한 노란색 반점들이며, 직경이 점차 증가하며, 이 반점들은 잎 밑표면에 주황색 uredinia (夏胞子堆)가 분화되기에 앞서 발생한다 (Figure 1). 심각한 녹병 감염 동안에는, 잎이 농포(pustules)로 덮이게 되어 조기에 떨어지게 하고 나무의 광합성 면적이 감소한다. 반복적인 감염은 나무를 약화시키고(debilitate) 가지가 말라버릴 수 있다 (branch dieback) [16,17].

H. vastatrix의 생리학적 균종들은 전 세계적으로 50가지 이상 확인되었으며 [1,3,18,19], 그 중 15개는 브라질에서 확인되었고, 브라질에서 우세한 균종은 II이다 [20-24]. H. vastatrix 생리학적 균종은 23가지 커피 차이 세트 (a set of 23 coffee differentials)의 독성 스펙트럼(spectra of virulence)에 따라 식별된다 [3,25,26]. 접종된 커피 잎 또는 잎 디스크의 농포(pustule)에 uredospores (하포자들)이 형성되면, 그 차이 클론(differential clone)은 취약한(susceptible) 것으로 분류된다 [27]. 이러한 차이 클론들(differential clones)은 CIFC에 의해 식별되어 다른 국가의 여러 연구 기관으로 보내졌다 [1,3,28].
CRL의 유행은 주로 온도와 습도의 영향을 받는다.
H. vastatrix의 Urediniospores (하포자들)는 잎이 6~24시간 동안 젖어 있을 때만 발아한다 [29].
21~25°C의 온도 범위와 빛이 없으면 발아가 촉진되는 반면 [30-32],
32.5°C 이상 및 12.5°C 이하의 온도에서는 발아가 억제된다 [33,34].
온도는 잠복기(latency period)에도 영향을 미친다. 일반적인 잠복기는 20~55일(보통 25~35일)이지만, 온도가 28°C보다 높거나 18°C보다 낮을 때 상당히 길어진다 [2,35]. 아침에 햇빛에 노출된 작물 쪽은 잎이 젖는 기간이 줄어들기 때문에 CLR 감염이 덜 나타난다. 반면, 커피나무의 아래쪽과 중간 1/3에 위치한 잎은 더 습한 미기후로 인해 바깥쪽 잎과 위쪽 1/3에 위치한 잎보다 감염 수준이 더 높다. 질병 강도가 더 높은 이러한 습한 환경은 밀도가 높은 재배 시스템, 그늘이 있거나 숲이 우거진 지역, 줄기가 많은 식물, 폐쇄된 작물이 많은 곳에서도 발생한다 [36].
CLR 발생률(incidence)과 심각도(severity)에 영향을 미치는 또 다른 측면은 나무의 영양 상태(plant nutritional status)이다. 발생률은 식물에서 병든 잎의 비율만을 추정하는 것과 관련된다. 대조적으로, 심각도는 병변 수 또는 기술적 규모를 기반으로 질병에 의해 영향을 받는 식물 조직의 영역(area of plant tissue affected by diseases)을 나타냅니다. 불균형한 영양 식물은 CLR의 영향을 더 많이 받고 더 큰 생리학적 마모(greater physiological wear)로 인해 더 취약해질 수 있게 된다 [17].
대부분의 아라비카 브라질 커피 재배지역에서는 주요 우기가 9월~10월 봄에 시작되어 12월~1월에 최고조에 도달한 다음, 6월~8월 사이 겨울에 최소 강수량으로 감소하기 시작한다 [37]. 온도 변동은 강수량과 유사한 패턴을 따른다 (Figure 2). 주요 개화 단계는 일반적으로 커피 나무의 최대 영양 성장이 시작되는 9월과 10월에 발생한다. 콩 충전(bean filling)은 1월부터 3월까지 이루어지며, 주요 수확기는 5월부터 7월까지이다.
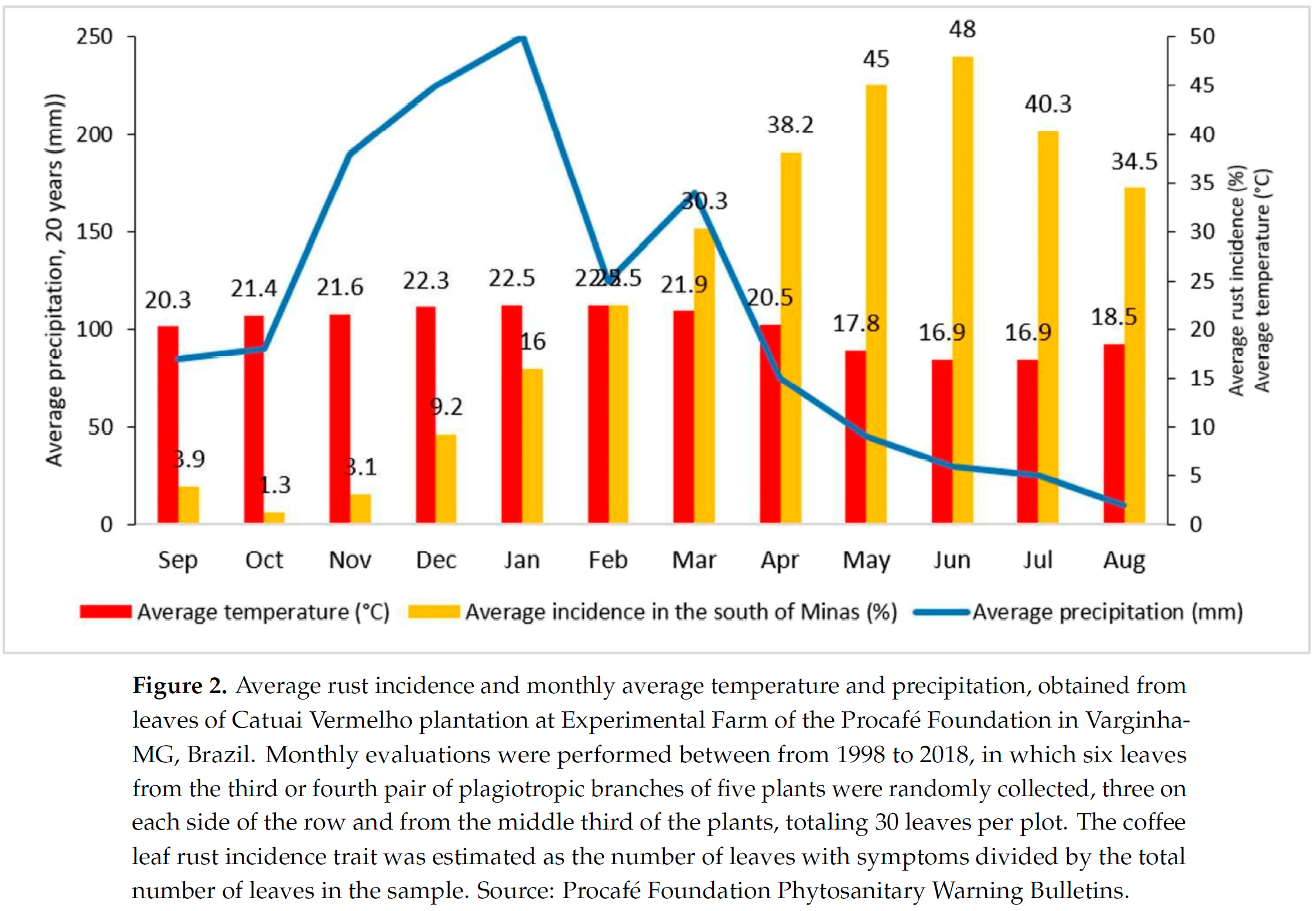
CLR 발병률은 11월 우기 이후부터 증가하기 시작하여 점차 증가하여 6월 늦가을에 최대에 도달한다. CLR 및 수확 작업으로 인한 피해로 인해 상당한 낙엽이 발생하여, 다음 우기가 시작될 때까지 커피 나무에 잎이 거의 남지 않는다. 9월부터 11월까지 나무 감염 수준은 감소하고, 11월~12월에 잔류 접종원이 새로운 주기를 다시 시작할 때까지 낮은 수준을 유지한다.
브라질 커피 재배의 성장 조건에 따르면, 커피 나무는 햇볕이 잘 드는 곳에서 자라며 수확량이 많고 수확량이 적은 해가 번갈아 가며 2년마다 열매 맺는 주기를 나타낸다. 해당 연도의 농작물 부하가 높든 낮든 상관없이 CLR 발생 곡선은 농작물 부하가 높은 해에 감염 수준이 더 높음에도 불구하고 동일한 행태를 보여준다 [1].

녹병에 강한 품종을 얻기 위한 커피의 유전적 육종은 1954년 브라질에서 Agronomic Institute (IAC)와 CIFC 간의 파트너십을 통해 시작되었다 [38].
CLR 저항성의 유전학에 대해 CIFC에서 수행된 선구적인 연구는 병원체의 생리학적 균종을 식별하고 국가 프로그램이 저항성 품종을 개발하는 데 도움이 되었다.
1970년대 녹병이 발생한 후 브라질의 기술 개발 및 커피 연구를 담당하는 다른 기관에서는 질병에 저항성을 갖는 새로운 품종 개발을 목표로 하는 유전 육종 프로그램을 시작했다.
⊙ Epamig (Agricultural Research Company of Minas Gerais)/
⊙ UFV(Federal University of Viçosa)/
⊙ UFLA (Federal University of Lavras),
⊙ Procafé Foundation (ex-IBC),
⊙ IDR-Paraná (Institute of Rural Development of Paraná, ex-Iapar), 그리고
⊙ Embrapa Café 등의 기관들이
모두 Research Café Consortium의 지원을 받았고,
Brazilian Agricultural Research Corporation (Embrapa Café)가 코디네이트하여 진행되었다.

CLR 저항성은 정성적/수직적(qualitative/vertical) 또는 정량적/수평적(quantitative/horizontal)일 수 있다.
정성적 저항성(질적 저항성, Qualitative resistance)은 SH 유전자라고 불리는 저항성에 큰 영향을 미치는 유전자 (주요 유전자 major genes)의 작용으로 인해 발생하며, 이는 동형접합성 상태(homozygous state)에서 높은 수준의 저항성을 촉진한다 [18,39,40].
정량적 유형의 저항성(양적 저항성, Quantitative-type resistance)은 중간 수준의 저항성을 촉진하는 2차 효과 유전자(secondary-effect genes) (minor genes)로 인해 발생한다 [40,41].
CLR과 커피의 상호작용에서 H. vastatrix의 생리학적 균종들은 커피 저항성 유전자들을 극복하려고 시도한다.
해당 균종에 해당 SH 유전자의 저항성을 파괴하는 병독성 유전자(v)가 없으면 포자형성 증상이 나타나지 않고 높은 저항성 반응이 발생한다. 각각의 SH 유전자의 저항성을 파괴할 수 있는 v 유전자(virulence gene)(신종)가 출현하더라도 식물은 여전히 중간 저항 수준을 촉진하는 minor 유전자들을 통해 녹병을 방어할 수 있다.
CLR에 대한 커피의 저항성 수준은 병원체에 대한 식물의 반응 유형, 증상, 및 질병 강도에 따라서,
⊙ highly resistant (HR),
⊙ moderately resistant (MR),
⊙ slightly resistant (SR),
⊙ susceptible (S), 또는
⊙ highly susceptible (HS)로 분류될 수 있다 (Table 1).
HR 및 SR levels을 가진 나무들은 일반적으로 다른 저자에 의해 각각 immune과 moderately susceptible로 불린다. HR level은 정성적 유형의 저항성 유전자 (qualitative-type resistance genes)에 의해 좌우된다. 그러나 이 수준으로 분류된 재배품종들은 일반적으로 정량적 저항성 유전자도 가지고 있다. MR, SR, S, 그리고 HS levels는 정량적 저항성(quantitative resistance)에 관계된다. HR level은 완전 저항성(complete resistance)이라고도 하며, MR 및 SR levels는 일반적으로 불완전, 중간 또는 부분 저항성(incomplete, intermediate, or partial resistance)이라고 한다.
Table 1. Description of CLR resistance levels and associated coffee cultivars.


CLR 저항성의 알려진 정성적 유전자로는 SH1, SH2, SH3, SH4, SH5, SH6, SH7, SH8, SH9 및 SH?이 있다 [18,25,42].
높은 질적 저항성을 촉진하기 위해 브라질에서 사용되는 주요 유전자 소스들은 Híbrido de Timor (HdT) 및 그 파생품종들 (예: Villa Sarchi × HdT, Caturra × HdT, Catuaí × HdT), Icatu 및 그 파생품종들 (예: Icatu × Catuaí, Icatu × Catimor, Icatu × Sarchimor), SH3 유전자와 그 파생물을 보유한 BA 시리즈의 커피나무 (예: Catuaí × BA-10), Geisha와 같은 재래종을 포함한 에티오피아산 야생 아라비카 커피나무이다.
최근 Barka et al. [43] 및 Almeida et al. [44]는 분자생물학 기술을 사용하여 HdT 832/1과 HdT 832/2에서 각각 SH10과 SH11로 명명된 두 개의 서로 다른 유전자를 발견했다.
255개의 H. vastatrix 분리주들이 2018년부터 2020년까지 브라질, 온두라스, 베네수엘라, 코스타리카에서 저항성을 잃은 HdT 파생품종들로 부터 수집되었다 [1]. 본 연구에서 병원형(pathotype)이라는 용어는 CIFC에서 사용된 방법론과 커피 감별 세트에 따라 균종으로 구분할 수 없는 H. vastatrix 분리주들을 지칭하기 위해 제안되었다. 10개의 H. vastatrix 분리주들 (Hv01~Hv10)이 브라질에서 확인되었으며, 그 중 3개는 Hv01 (v1, 5, 6, 7, 8, 9, and ?), Hv02 (v1, 5, 6, 8, 9, and ?) 그리고 Hv08 (v1, 2, 5, 6, 7, 8, 9, and ?)이다. 이들은 각각 HdT 832/1, HdT 832/2 또는 하이브리드 Kawisari 644/18을 감염시키지 않았고, 그들의 유전자형들은 게놈에서 더 많은 저항성 유전자를 가지고 있다는 것을 말한다. 10개의 브라질 병원형들을 구별하기 위해 20개의 구별 숙주들이 사용되었다; 그러나 그들은 분리주들을 균종들로 구별할 수 없었다. 그럼에도 불구하고, 병원체 게놈 인식은 병원체 분리주들과 differentiating hosts 사이의 상호작용에 의존하기 때문에, 이들 숙주는 10가지 병원형들을 구별하는 데 특히 중요했다.
HdT 교배에서 파생된 커피 나무는 SH5, SH6, SH7, SH8, SH9, and SH? 유전자를 개별적으로 또는 연합적 (in association)으로 포함하고 있기 때문에 브라질에서 CLR 질적 저항성의 중요한 원천이었다 [42]. 주요 브라질 HdT 커피 나무들은 HdT CIFC 832/1, HdT CIFC 832/2 및 HdT CIFC 2570이며 Caturra Vermelho CIFC 19/1 (Catimor 그룹 발생), Villa Sarchi CIFC 971/10 ( Sarchimor 그룹에서 유래) 및 Catuaí 그룹에서 재배한 품종 (Tables 2 and 3) 등이 있다. Catimor와 Sarchimor는 원래 교배의 자손 선발 뿐만 아니라 이러한 원래 교배의 후손과의 교배에서 직접 개발된 품종이라는 점은 주목할 만하다. 그러나 이들 품종 중 일부의 저항성은 나중에 아메리카 대륙의 분리주들과 병원형들에 의해 극복되고 공격을 받았다.

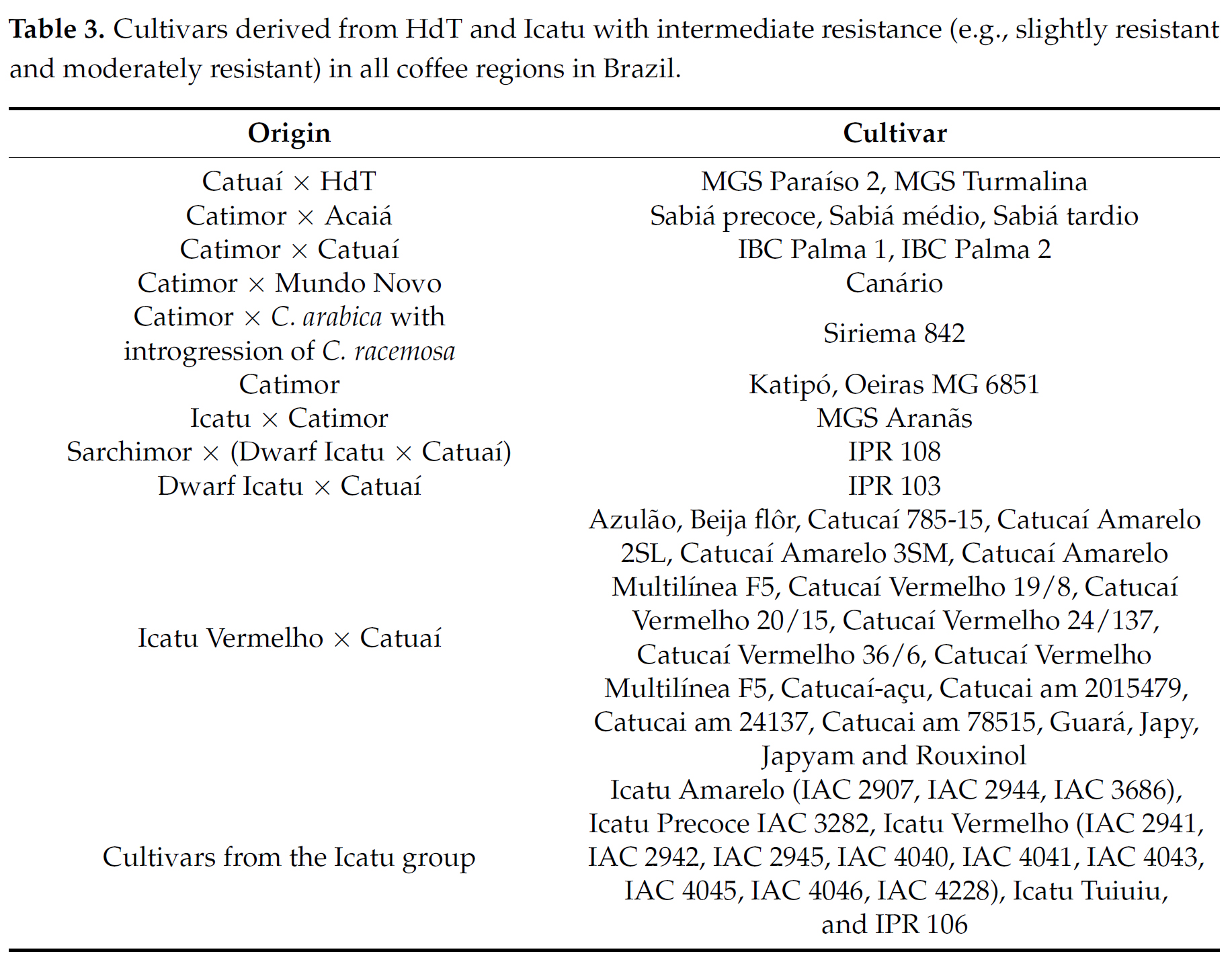
Icatu는 IAC에서 개발된 이중 염색체 종(doubled chromosome species)인 Coffea canephora var. Robusta, Coffea arabica var. Bourbon Vermelho 사이의 인공 교배에서, C. arabica Mundo Novo와 이 하이브리드의 역교배를 두 번 더 하여, 유래되었다 [45]. Icatu 외에도 IAC는 Mundo Novo와 Catuaí Amarelo를 교배한 "Dwarf Icatu"라는 커피 나무를 개발했다. 브라질에서 상업적으로 출시된 Icatu 그룹의 대부분의 품종은 이미 브라질 작물에 존재하는 H. vastatrix의 균종들에 의해 질적 저항성이 깨졌으며 품종 Icatu Amarelo IAC 2944 (Table 3)를 제외하고는 현재 SR 수준을 나타낸다.
Icatu Vermelho와 Catucaí라고 불리는 Catuaí 사이의 자생적 교배(spontaneous hybridization)로 인해 이미 CLR 질적 저항성이 손상된 여러 브라질 재배품종들이 생겨났으며, 이들 중 대부분은 현재 SR로 분류된다 (Table 3). Dwarf Icatu는 IAC에서 Catuaí와 교배되어 재배품종 IPR 102와 IPR 103을 생산했는데, 둘 다 정성적 저항성이 이미 깨졌다. 그러나 일부 지역에서는 IPR 102에 HR 수준이 있는데, 이는 IPR 103에 부족한 SH 유전자를 보유하고 있음을 나타낸다.
Icatu × Catuaí 그리고 Dwarf Icatu × Catuaí에서 파생된 커피 나무에서는 정성적 저항성이 이미 손상되었지만, 이러한 유전자형은 육종 프로그램에 사용할 수 있는 SH 유전자를 보유하고 있다.
SH1, SH2, SH4, 그리고 SH5 유전자들은 이미 여러 커피 지역에서 CLR로 대체되었으며(supplanted), Sarchimor 그룹의 재배품종들은 일반적으로 대부분의 커피 지역에서 높은 녹병 저항성을 나타낸다. 따라서 H. vastatrix 균종의 다양성으로 인해 일부 재배품종은 한 지역에서는 HR 수준을 나타내고 다른 지역에서는 SR 수준을 나타낸다. 예를 들어, 재배품종 Obatã (Sarchimor × Catuaí)는 Minas Gerais의 여러 장소에서는 HR을 나타내지만, Paraná 및 São Paulo 주의 일부 지역에서는 SR로 분류된다. 따라서 SH3는 브라질에서는 아직 파괴되지 않은 HdT 및 Icatu 커피 나무의 단일 저항성 요인들 중 하나이다. 에티오피아의 야생 아라비카 커피나무에 있는 SH1, SH2, SH4, SH5 유전자의 저항성은 브라질의 CLR에 의해 이미 파괴되었지만, 이러한 유전자형은 내구성 있는 저항성을 위한 유전자 피라미드에서 중요하다.
v3 병독성 유전자를 보유하는 H. vastatrix 의 생리학적 균종은 아직 브라질에서 검출되지 않았다 [26,46]. 따라서 SH3을 보유한 품종은 HR로 분류된다. SH3 캐리어인 BA 시리즈의 여러 커피 나무 중에서 IAC 1110이라고도 불리는 BA-10이 브라질에서 사용되는 주요 나무이다. BA-10 외에도 IAC 1111, IAC 1109, IAC 1112, IAC 1106, IAC 1116 및 IAC 1107로 각각 지정된 BA-2, BA-8, BA-13, BA-14, BA-16 및 BA-21과 같은 이 시리즈의 다른 커피 나무들도 사용된다.

커피나무의 정량적/수평적 저항성(quantitative/horizontal resistance)의 발현은 H. vastatrix 균종들이 SH 유전자들의 정성적/수직적 저항성(qualitative/vertical resistance)을 파괴하는 데 성공할 때 분명해진다. 마이너 유전자들 때문에 다양한 수준의 정량적 저항성이 관찰되었다. HdT 및 Icatu 나무들 [40]뿐만 아니라, C. arabica 및 C. canephora 품종들 [47,48]에서도 마이너 유전자들로 인한 불완전 또는 중간 저항성이 확인되었다. 이러한 마이너 유전자들에 의해 촉진되는 커피나무의 가장 일반적인 저항 수준은 SR과 MR이다. 그러나 Catuaí 그룹의 재배품종들과 같이 전 세계적으로 susceptible (S)한 것으로 알려진 커피나무들이라도 Bourbon 그룹의 재배품종들보다는 덜 취약하기 때문에 정량적 저항성을 촉진하는 마이너 유전자들을 가지고 있는 것으로 보인다. Bourbon 그룹과 같은 HS 재배품종들도 정량적 저항성 유전자를 갖고 있지만 질병 강도(disease intensity)가 크게 감소하지는 않는다. 발생률(incidence)과 심각도(severity)의 속성을 포함하는 "강도(intensity)"라는 용어는 여기에서 특정 에리어에서의 질병에 대한 일반적인 특성 측정 기준으로 사용되었다.
에티오피아 야생 커피나무들, HdT, Icatu, 그리고 그 파생물들도 SR 및 MR 수준에서 정량적 저항성의 중요한 원천이며, 브라질 BA 시리즈 커피나무들은, 그 자손들이 SH3 유전자를 상실하면 susceptible한 것으로 분류되므로, 정량적 저항성 유전자가 거의 없는 것으로 보인다. 야생 에티오피아 액세션들과 그 파생물들에서의 마이너 유전자들의 작용에 의해 유발되는 중간 저항성은 SH1과 SH4를 보유하는 나무들에서 더 흔하다. SH2 및 SH5를 보유하는 나무들은 질적 저항성이 손상될 때 일반적으로 취약하기 때문에 마이너 유전자들이 더 적은 것으로 보인다.

브라질에서 CLR 저항성 유전자들을 전달하기 위해 일반적으로 사용되는 육종 방법은 혈통(pedigree), 벌크(bulk), 그리고 역교배 (backcrosses)이다. 처음 두 가지 방법에서 초기 교배들은 서로 다른 질적 또는 양적 저항성 유전자들을 가진 부모 사이에서 발생하는 반면, 역교배는 하나 또는 두 개의 주요 유전자들을 전달하는 것을 목표로 한다. 단일 유전자형에서 질적 및 양적 SH 유전자의 피라미드화(pyramiding)는 CLR에 대한 내구성 있는 저항성을 갖는 재배품종 개발을 목표로 하는 개선 전략을 나타낸다. 정성적 유전자의 피라미드화는 혈통(pedigree), 벌크(bulk) 및 역교배(backcrosses)를 통해 수행될 수 있는 반면, 정량적 유전자는 처음 두 가지 방법을 사용한다.
주요 SH 유전자들과 관련된 분자 마커들의 개발이 이러한 유전자들을 피라미드화하여 육종 프로그램을 지원한다. SH3과 연관된 마커-활용 선발(Marker-assisted selection)은 이미 브라질의 육종 프로그램에 사용되고 있다. 마이너 저항성 유전자들과 연관된 분자 마커들의 추가 개발이, 이 유전자들의 미라미드화를, 심지어 역교배 방법을 통해서라도, 가능하게 할 것이다.
CLR이 브라질에 진출한 직후, Catimor, Sarchimor, 그리고 Icatu 그룹 뿐만 아니라, Catucaí라고 불리는 Icatu와 Catuaí 교배들로부터 저항성 재배품종 개발에 큰 중점을 두었다. 최초의 녹병 저항성 재배품종이 1980년대와 1990년대에 상업적 재배를 위해 출시되었다 [49]. 상업적 경작 직후에, 몇 가지 재배품종들, 주로 Catimor와 Icatu 그룹들, 그리고 이보다 덜한 정도로, Sarchimor 그룹의 재배품종들에서 저항성 중단이 나타나기 시작했다. 많은 경우, 저항성 와해는 불완전 그리고 부분적인 저항성이 남아 있었는데, 이는 아마도 마이너 유전자들의 작용으로 인한 것일 수 있다. 따라서 유전자 육종 프로그램은 여러 전략을 채택하여 이러한 저항성 와해를 극복하려고 노력해 왔다.
브라질 육종가들은 정성적 저항성보다 양적 저항성이 더 내구성이 있다는 점을 고려하여 다양한 마이너 유전자들을 결합한 품종 개발에 착수했으며 보다 내구성 있는 저항성을 목표로 삼았다. 바람직하게는 현재 육종 프로그램은 높은 내구성을 제공하고 CLR을 제어하기 위한 살균제 적용을 제거하거나 줄일 수 있기 때문에 정량적 저항성 유전자에서 유래하는 HR 및 MR 품종을 개발하는 것을 목표로 한다. HR 및 MR 품종은 SR과 SR, SR과 MR, MR과 MR 커피나무를 교배하여 마이너 유전자들로부터 개발되고 있다.
HdT 커피 나무와 그 파생물은 Icatu를 개발하는 데 사용된 나무와 다른 C. canephora 나무에서 유래되었다. 이러한 이유로 이 두 가지 저항성 소스의 조합은 마이너 유전자들의 수를 늘리고 MR과 같은 더 높은 저항성 수준을 갖는 나무를 생성하는 데 사용되었다. 에티오피아에서 온 야생 액세션들은 마찬가지로 Icatu, HdT 및 그 파생물과 교배되어 다양한 마이너 유전자들을 결합하고 저항성 수준을 높였다. 브라질에서는 이 목적으로 만들어진 다양한 교배 유형들이 Sarchimor × Icatu, Catimor × Icatu, Sarchimor ×(Icatu × Catuaí) 그리고 Sarchimor × 에티오피아산 야생 커피 나무이다. 일반적으로 Sarchimor, Icatu, Icatu × Catuaí 커피 나무와 에티오피아 야생 커피 나무의 질적 저항성 와해가 있을 때 저항성 수준은 HR에서 SR 및 MR로 감소한다.
커피 나무가 가지는 양적 저항성을 고려하지 않고 질적 저항성만을 기준으로 커피 나무를 선발할 경우, 마이너 유전자들의 유전적 침식(genetic erosion)이 발생할 수 있다. 정성적 저항성 와해를 경험하는 정량적 저항성 대립유전자 (alleles)가 거의 없는 재배품종은 녹병 저항성 수준의 감소로 인한 부분적인 저항성 또는 취약성만을 나타낸다. 예를 들어, Catimor 그룹에 속하는 재배품종 Oeiras는 1990년대 상업적 재배를 위해 출시되었을 때 HR 수준을 나타냈다; 그러나 몇 년 후 질적 저항력을 잃어 SR로 재분류되었다. SH2 및 SH5 유전자들을 보유하는 재배품종 IPR 100에서도 유사한 상황이 발생했다; 1980년대에 이 두 유전자들의 저항성 붕괴로 인해 품종 저항성은 Catuaí와 마찬가지로 S 수준으로 떨어졌다.
정성적 저항성 선발로 인한 이러한 정량적 저항성 손실을 피하기 위해 육종가는 주요 유전자와 마이너 유전자를 가진 커피를 교배하여 HR 커피 나무와 MR 식물과 같은 중간 저항성 식물을 선발해야 한다. 정성적인 저항성만을 고려하여, 브라질 육종가들은 HdT로 SH3을 운반하는 커피들과, SH5, SH6, SH7, SH8, SH9 및 SH? 유전자들을 운반하는 Sarchimor 사이의 교배들을 강화한다. 더욱이, 후자의 유전자형들은 SH1, SH2, 그리고 SH4 유전자들을 단독으로 또는 조합하여 보유하는 에티오피아의 야생 아라비카 커피나무들과도 교배되었다. 이러한 유전자형들을 사용하면 정성적 저항성이 와해되는 경우 중간 저항성을 목표로 하는 마이너 유전자들의 동시적 전달(simultaneous transfer)이 가능하다.

브라질에 등록된 138개 재배품종들 중 23개는 브라질 크롭들에 존재하는 생리학적 균종들로 대체된 주요 유전자들 (major genes)을 갖고 있지 않기 때문에 HR이다. 10개의 재배품종들이 일부 커피 지역에서는 HR이지만 다른 지역에서는 주요 유전자들이 대체되었기 때문에 MR 또는 SR이다; 그러나 그들의 마이너 유전자들은 다양한 수준의 중간 저항성 (intermediate resistance)을 촉진하는 역할을 한다. 일반적으로 브라질에서 여전히 HR인 브라질 재배품종은 Sarchimor, Catuaí × HdT 그룹들, 그리고 SH3를 보유하는 BA-10 파생품종들이다 (Table 2). HdT에서 파생된 재배품종들 중에서, 12개는 브라질의 모든 커피 지역에서 중간 저항성 수준(SR 및 MR)을 가지며(Table 3), MGS Aranãs 및 IPR 108은 HdT 및 Icatu의 주요 및 마이너 저항성 유전자들 나타낸다.
HdT CIFC 832/1과 Caturra Vermelho CIFC 19/1 사이의 교배로 Oeiras MG 6851과 Katipó라는 Catimor 그룹의 품종이 탄생했다. 이 재배품종은 SH 유전자를 포함하고 이전에는 HR이었지만 현재는 MR 및 SR이다. HdT CIFC 832/2와 Villa Sarchi CIFC 971/10 사이의 교배로 IAC 125 RN, IAPAR 59, IPR 97, IPR 98, IPR 104, Sarchimor MG 8840, IPR 99, 그리고 Tupi IAC 1669-33이라는 Sarchimor 그룹의 재배품종들이 탄생했는데, 이들은, 일부 지역들에서 MR과 SR 레벨을 나타내는 마지막 두개를 제외하고는, 브라질 전역에서 HR이다.
그 후 Sarchimor 그룹의 다른 재배품종들은 Sarchimor와 Catuaí 또는 Mundo Novo 사이의 인공적 또는 자발적인 교배를 통해 출시되었다; 이들은 Sarchimor보다 더 큰 생산 잠재력과 식물 활력을 나타내며 일부는 여전히 HR로 남아 있다.
Tupi IAC 1669-33 및 IPR 99와 같은 일부 Sarchimor 재배품종들이 브라질의 일부 지역에서 높은 저항성을 잃기 시작하더라도, Catuaí와 Mundo Novo와 같은 취약한 커피들과 교배된 Sarchimor 그룹의 재배품종들의 경우 질적 저항성 붕괴 가능성이 훨씬 더 높다.
재배품종 Obatã IAC 1669-20은 Sarchimor와 Catuaí 사이의 자발적 교배에서 파생되는 반면, Arara와 IAC Obatã 4739는 Obatã IAC 1669-20과 Catuaí Amarelo 사이의 자발적 교배에서 유래된다; 따라서 이들은 취약한 커피인 Catuaí와의 두 번의 교배를 보여준다. Tupi IAC 1669-33과 마찬가지로, 이 마지막 세 재배품종들은 브라질의 여러 지역에서 HR이지만 H. vastatrix의 더 치명적인 균종이 있는 일부 지역에서는 MR과 SR이다.
재배품종 Acauã, Acauãma, IPR Alvorada, Acauãnovo, Asabranca, Graúna, IPR 107, 그리고 IPR Pérola는 Sarchimor × Mundo Novo에서 유래되었다; 그러나 처음 3개는 여러 지역에서 HR이고, 다른 지역들에서는 MR 및 SR인 반면, 마지막 5개는 브라질 전역에서 HR이다. EPAMIG와 협력기관들은, Catuaí 그룹을 HdT CIFC 2570과 교배 시켜, Araponga MG1, Catiguá MG1, Catiguá MG2, MGS Ametista, MGS Catiguá 3, MGS Paraíso 2, MGS Turmalina, Paraíso MG H 419-1, Pau Brasil MG1, 그리고 Sacramento MG1 이라는 10개의 재배품종들을 개발했다. 일부 지역들에서 MR 및 SR인 Araponga MG 1과, 거의 모든 지역들에서 MR 및 SR인 MGS Paraíso 2 과 MGS Turmalina를 제외하고, 이들 모두는 브라질 전역에서 HR이다.
브라질에서는, Icatu 그룹에서 15개 재배품종들이 출시되었는데, IAC에서 13개, Procafé Foundation에서 1개, IDR-Paraná에서 1개가 출시되었다 (Table 3). MR인 Icatu Amarelo IAC 2944를 제외하고, 일반적으로, 모든 15개 Icatu 재배품종들은 CLR에 대해 SR 수준에서 중간 저항성을 나타낸다. MR 반응은 CLR에 유리한 환경 조건에 따라 최소한으로 달라지는데, 이는 아마도 이 재배품종이 SR 재배품종보다 저항성에 대한 마이너 유전자들을 더 많이 가지고 있기 때문일 것이다.
Icatu Vermelho와 Catuaí 사이의 자연 교배(spontaneous crossing)로부터, Procafé Foundation에서 19개 재배품종들이 탄생했다. 이들 재배품종들은 전형적으로 SR이고 때로는 S이다. 예외는 MR인 Azulão 재배품종이다. IPR 102 및 IPR 103이라고 불리는 IDR-Paraná에서 출시된 재배품종들은 Dwarf Icatu와 Catuaí 사이의 교배에서 유래되었으며, 각각 MR 및 SR 수준을 나타낸다. 그 현장에 존재하는 균종들에 따라, IPR 102는 일부 주요 유전자들이 아직 CLR에 의해 밀려나지 않았기 때문에, HR 재배품종으로서 행동할 수도 있을 것이다 (Table 3).
SH3-보유 커피 유전자형들로부터 파생된 세 가지 HR 브라질 재배품종들은 IAC Catuaí SH3, IPR 105, 그리고 Saíra이며, 각각 IAC, IDR-Paraná, 그리고 Procafé Foundation에 의해 등록되었다. 처음 두 개는 BA-10 (IAC 1110-8)에서 유래한 SH3를 갖고 있는 반면, 마지막 하나는 Catindu UFV 374 cv 643에서 유래했다. 재배품종 IPR 100은 BA-10에서 파생되었으며, SH3가 없기 때문에 녹에 취약하다 (Table 2). Mundo Novo와 Catuaí로부터 나온 재배품종들, 그리고 Catuaí × Mundo Novo groups로부터 나온 6개의 다른 재배품종들 (예: IAC Ouro Verde, MGS Epamig 1194, Maracatiá, MGS Travessia, Rubi MG 1192, Topázio MG 1190)은 S인 반면에, Bourbon으로부터 나온 재배품종들은 HS이다.

현재 브라질 커피 농장의 약 80%는 Catuaí 및 Mundo Novo 그룹의 CLR에 민감한 재배품종들로 구성되어 있다. 1990년대에 새로운 CLR 저항성 재배품종들이 도입되면서, 농촌 생산자들은 천천히 해당 재배품종들을 심기 시작했다. 1990년대와 2000년대 사이에는 Sarchimor 그룹의 HR 재배품종들에 대한 준수(compliance)가 거의 없었는데, 이들은 높은 생산 잠재력에도 불구하고 영양분과 물 공급 측면에서 부적절하게 관리되면 전형적으로 약한 식물 활력을 보인다. 2000년대부터 Sarchimor × Catuaí, Sarchimor × Mundo Novo, BA-10 파생종들, 그리고 Catuaí × HdT에서 유래된 더 나은 영양 활력을 지닌 새로운 CLR 저항성 HR 재배품종들이 출시되었다. 동시에, Icatu × Catuaí, Dwarf Icatu × Catuaí, 그리고 Icatu × Catimor에서 유래한 SR 및 MR 재배품종들이 출시되었다. 이들 새로운 재배품종들은 모두 CLR 저항성 외에도 더 강한 식물 활력과 높은 생산성, 우수한 컵 품질, 더 큰 열매 크기, Phoma spp.와 bacterial halo blight와 같은 질병에 대한 저항성, 선충류 Meloidogyne exigua, M. paranaensis 및 M. incognita에 대한 저항성, 가뭄에 대한 더 높은 저항성과 결합된 다른 바람직한 특성으로 인해 점점 더 많이 채택되는 왜성 재배품종들이다.
브라질 커피나무 부문에서 CLR 저항성 재배품종들의 비율이 낮음에도 불구하고, 지난 10년 동안 저항성 재배품종의 채택이 강화되었다. 그럼에도 불구하고 새로운 재배품종들은 일반적으로 생산자에게 추가적인 흥미로운 특성을 제공하기 위해 선택된다.

브라질에서 CLR의 화학적 방제는 50년 넘게 이 질병을 안고 생활하면서 단순화되고 합리화되어 효율성과 우수한 비용-효익 비율을 모두 보여준다. 따라서 올바른 살균제 선택, 사용량, 시간 및 살포 기술을 준수해야 한다. 일반적으로 관개 시설이 없는 지역에서 고급 기술을 사용하여 더 높은 수확량(이동 평균인 60kg 생 커피 30포/ha/ha 이상)을 얻기 위해 브라질 커피 농부들은 녹 발생률을 5% 미만으로 유지한다. 따라서 적절한 관리 기술이 필요하다 [50]. 이러한 기술 중에서 화학적 방제는 현재 사회적, 재정적, 환경적 지속 가능성 외에도 식량 안보를 보장하는 가장 좋은 방법이며, 방제는 일반적으로 농부가 이상적인 시간에 적절한 살균제 사용량을 적용하고, 숙주-병원체 관계 사이클을 방해하며, 그리고 질병 진행률을 감소시키는 것을 돕는 작물 모니터링, 의사 결정 지원 또는 예측 시스템을 기반으로 한다.
다양한 브라질 생물 군계는 다양한 농림토양기후 조건(edaphoclimatic conditions)과 지형 기복 형태를 갖고 있으므로 화학적 방제는 이러한 특성에 맞게 조정되어야 한다. 예를 들어, 가파른 경사가 있는 언덕이 많은 지역에서는 기계화 및 살포가 적용하기가 더 어려우므로 화학적 방제의 효율성이 떨어진다. 이러한 이유로 인해 CLR은 여전히 브라질 커피 재배 부문에서 문제가 되고 있으며, 녹 제거를 위해 이용 가능하고 효율적인 기술을 채택하지 않는 생산자에게 상당한 손실을 초래하고 있다. 살균제의 선택만큼 적용기술이 중요하다는 점은 주목할 만하다. 현재 브라질에서는 CLR의 화학적 방제를 위해 접촉식, 중간전신성 및 전신성 살균제 (contact, mesostemic, and systemic fungicides)가 이용 가능하다 [17].

Contact fungicides 접촉 살균제는 구리 및 dithiocarbamate 케미컬 그룹의 보호 살균제이다. 구리 화학 그룹 (cupric chemical group)에는 수산화구리(copper hydroxide), 옥시염화구리 (copper oxychloride), 황산구리(copper sulfate), 산화제1구리(cuprous oxide)와 같은 몇 가지 분자들이 포함되며, 이들은 여전히 다양한 제형으로 이용 가능하다 [51]. 처음 두 개는 브라질 커피 재배 부문에서 가장 일반적으로 사용되는 분자이다. 이러한 살균제들은 특히 작물 수확량이 적거나 생산성이 낮은 해에 CLR을 제어하는 데 도움이 될 뿐만 아니라 커피 재배에 사용되는 살균제의 활성 원리를 순환시키는 데 기여하여 전신 및 중간 살균제에 대한 저항성 병원균의 선택을 피할 수 있다 [2,17].
생산자들에게 제공되는 다양한 제품과 제제 중에서 가장 중요한 것 중 하나는 제제에 존재하는 금속 구리 또는 Cu2+의 양이다 [52]. 즉, 신진대사를 불가능하게 만들어 병원체를 죽일 수 있는 부위에 결합하려면 전하(charge) (이 경우에는 positive charge)가 있어야 한다. 이 정보는 일반적으로 살균제 패키지 인서트에 나와 있다. 곰팡이를 효율적으로 죽이려면 탱크 혼합물의 Cu2+ 농도가 1200ppm보다 커야 한다. 즉, 1000 L의 물이 담긴 현탁액에는 1.2kg의 활성 성분인 Cu2+가 포함되어야 한다.
접촉성 살균제나 보호성 살균제(protective fungicides)와 관련하여, 좋은 분사 장비를 유지하고 분사 기술에 투자하여 커피 잎의 양면을 잘 덮을 수 있도록 하는 것이 매우 중요하다. 특히 강우량이 여름철 내내 분산되는 브라질 조건에서 보호제 스프레이들 간의 간격을 추정하려면 보조제(adjuvants) 또는 제품 제제로 인한 접착이 필수적이다 [53].
또한, 적용 제품의 가용성에 따라 구리의 용해가 느려질 수 있다. 이는 식물이 다량의 금속 구리를 흡수하지 않도록 하여 식물 중독을 방지하는 데 중요하다. 살균제의 구리 농도는 1,200ppm 이상이지만 구리는 이 양만큼 식물에 흡수되지 않는다. 따라서 구리는 브라질 작물에서 자주 발견되며 순차적인 구리 스프레이와 잎의 농도는 150ppm(mg/kg) 이상이다. 이 농도는 식물 중독 증상을 유발할 수 있다. 그러므로,브라질에 심은 대부분의 재배품종들에서와 같이, 잎의 Cu2+ 농도는 이상적으로 70ppm (mg/kg) 미만으로 유지되어야 하며, 관개 되지 않은 지역에서는 헥타르 당 생두 60kg 봉지 30~35개를 생산하는 이동 평균이 필요하다.
구리 제품의 입자 크기는 접촉 면적에 반비례하기 때문에 고려해야 할 또 다른 변수이다. 그러나 이상적인 pH에서 침전물 없이 용량을 사용하여 나무 캐노피 또는 잎 영역의 가능한 최대 확장에 도달하는 잎 양면에 대한 효과적인 스프레이는 입자 크기를 보상할 수 있다. 대부분의 기존 상용 제품은 입자 크기가 다르며 다른 양으로 분산된다.
전통적인 크기의 구리 분자를 넘어서, 나노 입자들이 잎과의 접촉 면적이 증가하여 더 나은 적용 범위를 제공하므로 헥타르 당 적용되는 금속 구리의 양을 줄이는 데 기여할 수 있다. 이러한 감소는 여러 국가에서 농업에 금속 화합물이 미치는 영향을 최소화하기 위해 시행된 새로운 글로벌 법률에도 부합한다. 현재 나노 입자 농도는 250~500ppm(mg/kg)이며 크기 범위는 20~30nm인 100% Ag 및 Cu 나노입자가 있다.
특히 에틸렌디아민테트라아세트산 (ethylenediaminetetraacetic acid), 아미노산(amino acids), 질산염(nitrates)과 결합된 구리 킬레이트 (Copper chelates)는 다양한 제형으로 시중에서 구입할 수 있다. 대부분의 경우 금속 구리의 농도는 300 ppm(mg/kg) 미만이므로 그 효과는 영양과 저항을 모두 유발한다. 따라서 이러한 킬레이트는 아데노신트리포스페이트 (adenosinetriphosphate) 및 아데노신디포스페이트 (adenosinediphosphate) (에너지), 아미노산 및 미량 영양소와 같은 화합물 뿐만 아니라, 리그닌 (lignin)과 페놀들(phenols)와 같은 저항 산물을 만들어내는 시킴산 (shikimic acid) 및 자스몬산 (jasmonic acid) 경로의 산물을 만들어내기 위해서는 양호한 영양 상태와 조건을 가진 나무에 두 번 이상 적용되어야 한다. 킬레이트는 제제에 따라 병원체에 직접적인 독성(direct toxicity)을 거의 나타내지 않는다. 어떤 것들은 개화기 동안 살포해야 하는 반면, 살균제와 혼합된 다른 제형들은 브라질의 우기, 즉 열대 여름 동안 적용해야 한다. 이러한 킬레이트는 높은 흡수성을 나타내어 Cu2+를 쉽게 이용할 수 있게 해준다. 그러므로 식물독성(phytotoxicity)을 피하기 위해 다량의 요소를 제공해서는 안 된다.
Mancozeb은 dithiocarbamate 화학 그룹에 속하는 보호 살균제(protective fungicide)이다.
이는 또한 트리아졸 (triazoles), 카르복사미드 (carboxamides) 및 스트로빌루린(strobilurin) 살균제와 결합하여 곰팡이 저항성 관리에 사용되는 다중 현장 살균제(multisite fungicide)이다. 그 제제에는 커피나무 영양에 기여할 수 있는 Zn과 Mn도 포함되어 있다. 더욱이, 트리아졸 및 스트로빌루린을 함유한 삼중 혼합물이 이미 시판되고 있다.
접촉형 살균제의 살포는 잎의 배축면(abaxial surface, lower)과 향축면(adaxial surface, upper)을 모두 덮도록 노력하 여 효과적으로 수행되어야 한다. H. vastatrix는 잎의 밑면에 있는 기공을 통해 침투한다. 따라서 병원균 침투를 방지하기 위해 제품으로 철저히 덮어야 한다. 다 자란 커피나무의 살포량은 나무 크기, 낙엽, 그리고 식재 및 재배 밀도에 따라 최소 200L ha−1(나무 높이 최대 1.5m까지), 최대 500L ha−1(나무 높이 3.5~4m)이어야 한다. 브라질에서 개발된 재배품종들은 다양한 biomes에서 재배되기 때문에, 캐노피 구조, 노드 간 길이, 나무 높이, 잎 크기 및 경사굴성 가지 강도(intensity of plagiotropic branching)가 다르기 때문에, 살포 효율성을 방해할 수 있다. 따라서 시럽의 양을 고려하는 것이 특히 중요하다. 더 큰 캐노피 직경, 닫힌 캐노피 구조 및 경사굴성 가지의 더 높은 분지 강도(higher ramification intensity)를 가진 키가 큰 나무들에는 더 많은 시럽량이 필요하다. 또한, 제품이 잎 양면에 균일하게 닿을 수 있도록 권장량에 따라 정확한 시럽 량을 도포할 수 있도록 스프레이 노즐의 상태가 양호해야 한다.

Mesostemic fungicide 그룹에는 다음과 같은 분자들을 가진 이 포함된 스트로빌루린(strobilurins)이 포함된다:
azoxystrobin,
metominostrobin,
picoxystrobin,
pyraclostrobin 및
trifloxystrobin.
이는 일반적으로 트리아졸(triazoles)이나 카르복사미드(carboxamides) 또는 둘 다와 혼합되어 전신 살균제와 mesostemic 살균제의 혼합물을 생성한다 (MixSM).
더 나은 효과를 위해서는 미네랄 및 식물성 오일과 같은 보조제와 함께 사용하는 것이 좋은데, 이들은 왁스 층을 감소시키고 잎, 꽃, 가지 및 과일의 표면 큐티클의 표면 장력을 파괴하고 병원체에 대한 치사량을 흡수를 제공할 수 있다.

System fungicides 전신 살균제에는 브라질의 CLR 및 기타 커피 병원균을 방제하기 위한 트리아졸 (triazoles), 스트로비룰린 (strobirulin), 카르복스아미드 (carboxamides )의 화학 그룹이 포함된다 [17].
알파벳 순서로, 현재 사용되는 트리아졸 분자들에는
시프로코나졸 (cyproconazole),
에폭시코나졸 (epoxiconazole),
플루트리아폴 (flutriafol),
테부코나졸(tebuconazole), 그리고
트리아디메놀(triadimenol)이 포함되는 반면,
카르복스아미드 분자(carboxamide molecules)에는
벤조빈디플루피르(benzovindiflupir),
플럭사피록사드(fluxapyroxade) 및
티플루자마이드(thifluzamide) 등이 있다.
이러한 살균제는 엽면 적용을 통해 살포할 수 있으며 스트로빌루린과 병용하는 것도 권장된다; 접촉 제품과의 삼중 혼합물도 이 제제에 존재한다.
토양에서 트리아졸(triazoles)은 기술 권장 사항에 따라 10월과 11월 사이 우기가 시작될 때 나무 캐노피 아래에 물을 뿌리거나 분출하여 단독으로 또는 살충제와 혼합하여 적용한다. 살포하기에 이상적인 순간은 토양이 촉촉하고 뿌리를 통해 이러한 생성물을 더 잘 흡수할 수 있는 때이기 때문에, 다양한 농림토양기후적 조건과 생물군계에서 기후를 모니터링하는 것이 중요하다. 현재 브라질의 고-생산성 시스템에서 CLR을 제어하려면 이 토양 적용 방법을 엽면 살포와 결합해야 한다. 또한 생산자와 기술자는 다양한 재배 시스템에서 다양한 품종의 나무 캐노피에 있는 가지와 잎의 수에 따라 과소 또는 과다 투여를 방지하고 비효율적인 처리 또는 식물 독성을 방지하기 위해 항상 나무 크기를 알고 있어야 한다.
스트로빌루린과 혼합된 트리아졸의 효율성은 건조지에서 60kg 짜리 30 bags 이상, 관개 농작물에서 헥타르당 그린 커피 60kg 짜리 40 bags를 초과하는 이동 평균을 갖는 생산 시스템에서 접촉 살균제의 효율보다 의심의 여지 없이 더 높다. 그러나 접촉성 살균제는 트리아졸 및 카르복스아미드와 같은 특정 작용 방식을 갖는 전신 살균제에 대한 병원체 저항성을 관리하는 다중 현장 살균제로서 필수적이다. 더욱이, 접촉성 살균제는 병원체 집락화 단계에서 보다 효율적인 전신치료효과 (systemic curative effects)를 나타내고 포자 형성 또는 번식 단계에서는 박멸 효과 (eradicative effects)를 나타낸다.
전신 살균제는 잎 내에서 매우 효율적으로 분포되지만, 살포는 접촉 살균제만큼 주의해야 한다. 상대습도가 낮거나 (40% 이하) 온도가 높을 (30°C 이상) 가능성이 있는 경우에는 분사 효율이 저하되므로 환경 조건이 개선될 때까지 분사를 중지해야 한다. 즉, 살포는 가능하면 이슬이 많은 아침 이른 시간에 하거나, 하루 중 가장 더운 시간을 피하여 늦은 오후와 이른 저녁에 하는 것이 좋다.
전신 살균제에 영향을 미치는 다른 두 가지 중요한 요소는 살포 시럽의 수질과 터보 분무기 및 배낭 분무기와 같은 살포 장비의 상태이다. 물은 뿌리는 시럽과 유사하게 pH 6~7.5 사이에서 깨끗하고 점토가 없어야 한다. 트리아졸과 카르복스아미드는 흡수와 전신 작용에 영향을 미치는 극성 하전 화합물(polar charged compounds)이다. 따라서 이러한 특성을 손상시킬 수 있는 제품을 탱크에 혼합하는 것은 피해야 한다. 또한, MixSM 엽면 살포(foliar applications)를 함께 혼합하는 것보다 미량 영양소 및 구리 엽면 살포를 산재시키면(interspersed) 방제 효율성이 더 높아진다.

일부 저자들은 질병 삼각관계의 변수, 즉 병원체, 숙주, 환경을 기반으로 살포 계획을 세우는 것을 목표로 질병의 발병을 예측하는 모델을 제안한다 [54]. 이러한 모델은 주로 CLR 심각도 값 (severity values), 유리한 잎 온도 및 습도 발생, 기상 데이터를 기반으로 한다.
1960년대 이래로 질병 발병의 다양한 지표를 예측하고 질병 관리에 도움이 될 수 있는 20개 이상의 모델들이 개발되었다 [55-59]. 그러나 이러한 모델은 아직 커피 농장의 녹병을 방제하는 데 사용되지 않는다. 브라질의 여러 커피 지역에는 온도, 강수량, 커피 농장의 CLR 발생과 같은 기후에 대한 데이터를 공개하는 연구 기관에서 개발한 경고 시스템이 있다.
브라질 커피 작물의 기후, 식재 및 관리 여건을 고려하여 동 제품(cupric products)만을 적용하는 방제방법이라면, 우기 시작부터 끝까지 예방적으로 적용하고 강우량에 따라 20~30일 간격으로 반복 적용해야 한다. 그러나 브라질에서는 구리 살균제를 독특하게 사용하는 경우가 드물고 식물 독성을 유발할 수 있다. 일반적으로 이러한 살균제는 스트로빌루린을 분사하여 트리아졸이나 카르복사미드와 혼합하거나 사이 사이 섞어 사용할 수 있는(interspersed) 다중 현장 제품(multisite products)이기 때문에 저항성 관리를 위해 사용된다.
CLR을 방제하기 위해 브라질 커피 작물에 적용되는 가장 일반적인 살균제는 전신 및 mesostemic 또는 MixSM으로, 살포 횟수는 45~60일 간격으로 2~4회이다. 이러한 간격과 적용 횟수는 기상 조건, 특히 비와 유리한 온도, 식물 영양, 식재 간격, 생산성, 품종의 저항 수준 및 가지치기 또는 흡지 제거와 같은 기타 재배 관행에 따라 달라진다 [60]. 화학적 방제의 시작은 CLR 증상을 나타내는 잎의 비율을 기준으로 하지 않는다. 취약성 재배품종들에 따라 생산성이 높은 브라질 작물의 경우 증상이 나타난 후 살균제를 적용하면 방제가 덜 효율적이라는 사실이 이전 연구에서 밝혀졌기 때문이다. 이는 CLR이 이미 다른 잎을 감염시켰지만 증상이 나타나지 않아 제어가 늦어져 효율성이 떨어지기 때문이다. 따라서 브라질에서 CLR을 통제하기 위한 엽면 살포는 일반적으로 증상이 나타나기 시작하는 12월에 시작된다. 이 기간은 강수량과 기온의 증가와 일치하며 (Figure 1), 이는 CLR 확산에 유리하다. 따라서 적용은 일반적으로 12월부터 4월까지 수행되며, MixSM을 3회 스프레이하거나 구리 살균제만 5회 스프레이한다. 대부분의 경우 MixSM은 보호 살균제와 결합된다. 그러나 MixSM에 잎의 미량 영양소가 포함된 보호 살균제 스프레이를 두 번 더 뿌리면 방제가 더 효율적이다.
많은 브라질 작물은 생산성이 매우 높으며 Catuaí 및 Mundo Novo 그룹과 같이 CLR에 취약한 재배품종들의 이동 평균은 헥타르당 생두 50~60kg bags 이상에 달한다. 이러한 소출의 경우, 취약한 재배품종들에는 최대 4번의 전신, mesostemic 또는 MixSM 스프레이가 필요할 수 있다. 특히 브라질의 수확기인 가을과 겨울의 높은 강수량과 아주 비슷하게 발생하는 정상 이상의 강수량이 있는 여름의 경우처럼, 환경 조건들이 곰팡이 증식에 좋은 경우라면 더욱 그러하다.
트리아졸을 단일 적용하여 토양에 살균제를 살포하는 경우, 효과적인 CLR 방제를 위해 MixSM을 2~3회 엽면 살포하는 것으로 보완할 필요가 있다. 토양을 통한 살포는 엽면 살포 전에 수행된다. CLR 관리에 기여하는 것 외에도, 토양 적용은 해충을 방제할 수도 있다. 왜냐하면 살균제가 살충제와 혼합되어 뿌리 시스템과 커피나무 크라운의 활력을 증가시키기 때문이다.
Icatu, Icatu × Catuaí group 그리고 일부 Sarchimors derivatives와 같이 SR 및 MR 수준에서 CLR에 대한 중간 저항성을 갖는 재배품종들의 경우, 일반적으로 취약성 재배품종들에 필요한 것보다 1~2회 적은 스프레이로 CLR을 방제하는 것이 가능하다. 살포 횟수의 감소는 수확량 목표, 토양 비옥도 및 기상 조건에 따라 달라진다. 낮은 소출과 같은 일부 상황에서는 중간 정도의 저항성 재배품종에 대해 CLR의 화학적 방제가 필요하지 않다.
그러나 CLR을 방제하기 위해 HR 재배품종을 사용하는 재배자라도 Cercospora coffeicola로 인한 갈색 눈 반점병 (brown eye spot)과 같은 다른 질병을 방제하기 위해서는 살균제의 사용이 여전히 필요하다. 후자는 CLR과 거의 동시에 발생하며 이러한 질병을 통제하기 위해 사용되는 살균제는 거의 항상 동일하다. 따라서 이 경우에는 HR 재배품종이 있는 작물에도 살포된다. brown eye spot은 때때로 CLR보다 덜 공격적인 질병이므로 더 적은 수의 스프레이가 필요하다. 따라서 HR 재배품종이 있는 작물에 살균제 사용이 필요하더라도 필요한 적용 횟수는 일반적으로 S 및 HS 품종에 비해 적다.
CLR을 방제하기 위한 HR 재배품종을 포함하는 작물에서 생산성은 헥타르당 60kg 그린 커피 40 bags 보다 높으며, 생산자는 여름 동안 종종 2~3회 살포를 계속한다. CLR을 방제하기 위해 HR 재배품종을 포함하는 작물에 채택된 관리, 기후 및 재배 실무에 따라 brown eye spot의 강도가 감소할 수 있으며 이로 인해 MixSM 적용 횟수가 줄어들 수 있다.
브라질의 커피 재배조건은 숲과 Cerrado를 보존하기 위해, 높은 면적당 생산성을 요구하며, 이로 인해 질병에 취약한 재배품종들에 화학적 방제가 사용된다. 예를 들어, 브라질 최대의 커피 생산지인 Minas Gerais에서는 평균 34%의 부지가 합법적인 산림과 보호구역을 영구적으로 보호하도록 지정되어 있다. 화학적 방제와 관련하여, 기술 및 농경 학교에서 교육을 받은 자격을 갖춘 기술자는 대부분의 시설에서 정확한 복용량, 시간 및 스프레이 빈도에 따라 적절한 살균제를 적용할 것을 권장한다. 따라서 살균제는 병원체에게는 독이 되고 식물에게는 약이 될 뿐이므로 농약 잔류물이 없는 커피 생산으로 식량 안보를 보장할 수 있다.

커피 경작 시행 및 관리에서, 올바른 재배 실무의 채택은 질병 감소를 조절하고 화학적 방제를 용이하게 하여 CLR을 통제하는 데 도움이 될 수 있다. 강수량, 온도, 커피 가지들에서의 햇빛 위치,수형과 같은 비생물적 환경 요인은 CLR과 천적 개체군의 역학에 중요한 역할을 한다 [2,12,61-63]. 커피 플랜테이션 형성에서는, 간격, 재배품종, 영양, 그리고 잡초 관리의 선택이 중요하다 [64]. 질병 관리를 용이하게 하기 위해 작물 특성도 고려해야 하며, 작물 수량, 토지 지형, 재배 시스템의 크기 및 유형, 생산자의 기술 수준, 작업에 사용할 수 있는 장비 및 노동력, 물 가용성, 각 지역의 기후 조건, 다른 해충 및 질병의 동시 발생, 그리고 그 시스템의 비용/효익도 중시해야 한다.
더 조밀한 간격을 사용하면 축축하고 그늘진 환경이 조성되어 CLR 감염이 촉진된다 [65]. 브라질 커피 재배지역에서 흔히 볼 수 있는 기계화된 줄에 심는 방식은 줄 사이는 더 많이 개방되고 줄 내에는 덜 개방된 것이 특징이며, 통풍이 잘 되고 작물 내 습윤 기간이 짧아진다. 또한 트랙터와 분무기의 통과를 용이하게 하고 연속적인 식물 열에서 분무 용액 낭비를 줄인다. 현재 브라질 시장에서 사용할 수 있는 장비를 사용하면 살포된 방울로 더 쉽게 도달가능한 작은 재배품종이 더 낫다. 심은 후 새싹들(sprouts)을 가지 치기하지 않아 줄기가 과도하게 늘어나도 촉촉하고 그늘진 환경이 조성된다. 따라서 필요할 때마다 커피나무를 적절히 솎아내고 가지치기를 하여 나무의 높이를 줄이고 커피 캐노피의 통기성을 개선해야 한다. 덜 습한 미기후를 확보하고, 잎이 젖어 있는 기간을 줄이기 위해서는 흡지들과 과도한 가지들을 제거하는 것도 중요하다. 게다가, 매우 키가 큰 잡초는, 작물, 특히 어린 작물에 수분을 축적할 뿐만 아니라 성체 나무 캐노피의 하부 1/3에 그늘을 만들고 감염을 촉진한다.
잡초는 또한 토양에 적용된 살균제를 흡수하여 커피나무에 대한 가용성을 감소시킬 수 있다. 효과적이고 균형 잡힌 양분 수준을 제공하는 적절한 시비는 나무가 질병에 덜 취약하도록 만들고, H. vastatrix 감염 후 회복을 촉진한다.
요약하면, 통합적 커피 녹병 관리(integrated coffee rust management)를 제안할 때에는 이러한 모든 요소를 고려해야 한다.

브라질에서의 CLR 도달은 처음에는 브라질 커피 생산에 심각한 위협으로 여겨졌다. 그럼에도 불구하고, 이 문제를 해결하기 위해 취한 조치는 커피 재배 부문의 리뉴얼과 현대화로 이어졌으며, 커피 산업을 더욱 확립하고 브라질의 커피 생산량을 증가시켰다.
최근 수십 년 동안 브라질의 유전적 커피 육종 프로그램은 CLR에 저항성을 갖는 많은 재배품종들을 개발했다. 그럼에도 불구하고, 커피 재배자들이 이러한 새로운 재배품종들을 채택하는 속도는 더디었다. 전통적이고 취약성이 있는 재배품종들의 성과가 좋았고, 그리고 화학적 녹병 방제방법은 비용이 저렴하고 효율성이 높기 때문이다.
또한 커피 재배자와 기술지원 담당자에게 새로운 재배품종을 전파하기 위한 전략도 필요하다. CLR에 대한 저항성은 브라질 커피 재배자들의 재배품종 선택을 결정하는 유일한 특성은 아니다. 높은 소출, 콩의 사이즈, 나무의 활력, 가지치기에 대한 반응, 성숙주기, 쉬운 기계 수확, 다른 질병들과 선충류 등에 대한 저항성 등의 특성들도 재배품종 선택 시 중요하게 고려된다. 이러하여, 브라질의 대부분 커피 플랜테이션들은, Catuaí와 Mundo Novo와 같이 여전히 CLR에 취약하다.
CLR 관리에는 화학적 방제의 사용이 일반적으로 사용되지만, 재배 관리의 활용, 농약에 대한 안전 실무의 사용 및 살포 횟수를 최소화하기 위한 예측 모델을 활용하면 살균제의 합리적인 사용이 가능하다. 따라서 환경적 지속가능성 (environmental sustainability)은 농업 생산의 경제적, 사회적 요구와 균형을 이룰 수 있다.
브라질 커피 생산의 상당 부분이 기계화가 불가능한 산악 지역에 위치하여 녹병에 대한 화학적 제어가 어렵고 상당한 생산 손실이 발생한다는 점이 눈에 띈다. 이러한 지역에서는 저항성 재배품종들이 CLR을 제어하는 가장 효율적인 방법이며, 느리지만 확실하게 전통적인 재배품종들을 대체하고 있다.
CLR은 상당한 경제적 어려움을 초래할 가능성이 있는 이 나라에서 가장 중요한 커피 질병임에도 불구하고, 브라질 커피 재배자들은 이 질병과 함께 살아가는 법을 배워오고 있다.




'Brazil Coffee' 카테고리의 다른 글
| 브라질 커피 서스테이너블리티, 생산, 인증 (0) | 2023.04.21 |
|---|---|
| 브라질 커피 생두 등급 분류를 위한 품질기술규정 (0) | 2023.04.16 |
| 브라질의 커피 원산지 표기 (0) | 2023.04.16 |
| 브라질 커피 산지 (Coffee Regions - Indicação Geográfica) (0) | 2023.04.11 |
| 브라질 커피 역사 (0) | 2023.04.09 |




댓글