
세계에서 가장 중요한 무역 상품 중 하나인 커피는 50개 이상의 열대 개발도상국에서 재배되고 있으며, 최소 1억 명에게 중요한 소득원과 고용원을 제공한다 (Vega, Rosenquist & Collins, 2003).
Coffea arabica L.과 C. canephora Pierre ex A. Froehner의 두 종이 전 세계 커피의 약 99%를 생산한다.
이들은 Coffea 아속(Rubiaceae과)에 속하며, 여기에는 아프리카(41종), 마다가스카르(51종) 및 마스카렌(3종)의 열대림 고유 야생종도 포함된다 (Davis et al., 2006).
지난 20년 동안 이 屬의 분류학은 근본적으로 개정되었고 많은 새로운 종이 기술되었다
(Bridson & Verdcourt, 1988; Stoffelen et al., 1997; Stoffelen, 1998;
Stoffelen, Robbrecht & Smets, 1999; Davis & Rakotonasolo, 2000; Davis, 2001;
Davis & Rakotonasolo, 2001a, b; Cheek, Csiba & Bridson, 2002;
Davis & Rakotonasolo, 2003; Davis & Mvungi, 2004; Sonké & Stoffelen, 2004).
카메룬, 탄자니아, 마다가스카르는 가장 많은 수의 Coffea 종을 보유한 국가이다.
육종 관점에서 야생종은
┌ 질병 및 해충 저항성,
├ 특정 환경 조건에 대한 내성,
└ 커피 제품의 품질과 관련된 생화학적 특성과 같은 중요한 특성의 원천이다 (Anthony, Astorga & Berthaud, 1999).
1966년부터 1987년까지 살아있는 커피나무는
마다가스카르, 코모로, 마스카렌 섬과
아프리카 8개국 (Guinea, Ivory Coast, Cameroon, Central Africa, Congo, Ethiopia, Kenya and Tanzania)에서 Bioversity International (구 IPGRI),
파리 자연사 박물관 (the Paris Museum of Natural History, MHNP),
프랑스 국제개발농업연구센터 (the French Agricultural Research Centre for International Development, CIRAD)와 협력하여
the Institute of Research for Development (IRD, 몽펠리에, 프랑스)에 의해 수집되었다
(Anthony, Dussert & Dulloo, 2007).
대규모 필드 유전자 은행이 여러 열대 국가, 특히 Ivory Coast (아프리카 Coffea 종), Ethiopia (C. arabica) 및 마다가스카르 (토종 Coffea 종)에 설립되었다.
Cameroon의 바코시 삼림보호구역 (the Bakossi Forest Reserve)에서 코페아 표본의 cuttings를 수집했고 (Anthony, Couturon & de Namur, 1985), Ivory Coast의 Man-IRD coffea 컬렉션에서 재배했다 (Anthony, 1992).
수집가들은 이 컬렉션을 기술된 코페아 종 중 어느 것에도 귀속시킬 수 없었다.
문제의 종은 1997년까지 분류학 연구자들에게 연구되지 않은 채로 남아 있었는데, 그 때 이 종의 표본 자료가 준비되어 IRD에 보관되었다.
이 표본 자료에 대한 형태학적 연구와 BR, BRLU, K, P 및 WAG의 코페아 자료에 대한 재검토 후,
이 표본이 새로운 종을 나타낸다는 것이 확인되었다.
이 종의 다른 컬렉션은 알려져 있지 않다.
형태학적 및 생화학적 데이터를 사용하여 인식에 대해 논의한다.
- 식물 표본 약어는 Holmgren, Holmgren & Barnett(1990)을 따른다.
- 형태학적 측정 및 기술은
BR, BRLU, K, P 및 WAG (콩고-브라자빌, 콩고-킨샤사, 카메룬 및 가봉에서 가장 중요한 식물 표본)의 식물 표본을 기반으로 한다. - 컬렉터 번호가 없는 표본은 고유한 ID를 제공하기 위해 고유한 BR 표본 바코드로 참조되었다 (예: BR-S.P. 872411).
- 종자 껍질은 광학 현미경으로 표면에서 관찰했다.
- 피렌 (pyrenes)을 끓인 후, 종자 껍질을 분리하고 호이어 검 (Hoyer gum)으로 투명한 영구 제제를 만들었다.
- 해부학적 관찰을 위한 잎 샘플은 몽펠리에의 IRD 컬렉션에서 수집했다.
- 샘플은 개미산 (formic acid)에 보존하여 BR에서 보관하고 거기에 보관했다.
- 잎의 횡단 슬라이드(Transverse slides)를 만들고, 임계점(critical point)을 말리고 금으로 코팅했다.
- 해부학적 관찰은 주사 전자 현미경(Jeol 5800 LV)으로 수행했다.
- 이 논문에서 우리는 꽃차례의 컵 모양 구조에 대해 calyculi라는 용어를 사용한다
(자세한 논의는 Davis, Bridson & Rakotonasolo, 2005 참조).
Diagnosis :
propter foliis minoribus, corollae tubo minore et antheribus minoribus differt ab speciibus
Africanae Centralis.
(더 작은 잎, 더 작은 화관통 및 더 작은 꽃밥 때문에 중앙아프리카 종과 다르다.)
Type :
- Anthony 17 (BR!, holotype (정기준표본); 바코드 BR-S.P. 000000872411이 있는 표본),
1997년 Man-IRD 커피 컬렉션, Ivory Coast에서 수집. - 카메룬, 남서부 지방, Tombel Division, Bakossi Forest Reserve (바코시 삼림 보호 구역),
Mungo River의 다리 동쪽 2km, N4°44′ E9°35′, 고도 160m, 1983년 2월 14일에 수집한 cuttings에서 재배.
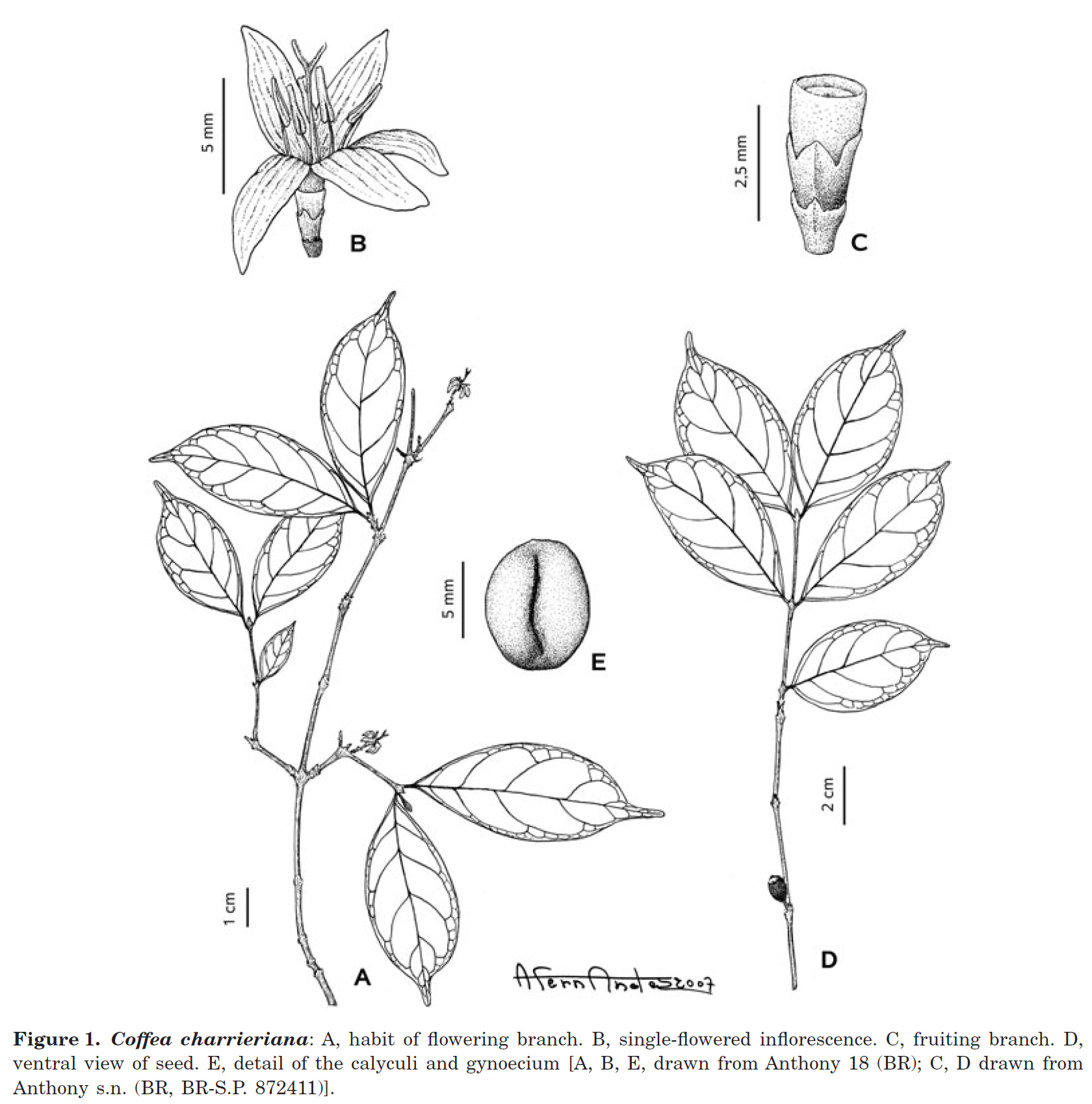
Description :
- 樹高 1~1.5m의 灌木 (Shrub).
- 작은 가지(Branchlets) → 지름 1~2mm, 軟毛(柔毛)가 있음(pubescent).
- 턱잎 (托葉, Stipules)
→ 삼각형~삼각주 모양(deltate), 약 2mm 길이, 인편 모양(imbricate),
→ 연모가 있고 가장자리에 털이 있음. - 잎자루 (Leaves petioles, 葉柄) → 길이 약 2mm;
- 잎새 (leaf blades)
→ 타원형(elliptic) 또는 넓은 타원형(드물게 도란형 obovate),
→ (4–)5–7(–8) × 2.2–3.5cm, 얇음(thin); - 잎밑 (leaf base, 葉底) → 銳底(acuminate)에서 약간 流底(slightly attenuate);
- 잎끝 (apex, 葉先) → 끝이 둥글고(rounded tip) 뾰족하며(acuminate), acumen은 → 길이 7–13mm,
- 잎면 (leaf surface) → 위아래로 털이 없음(glabrous);
- 2차 맥 (secondary veins)은 → 주맥(midrib) 양쪽에 5–7개;
- 3차 맥 (tertiary veins)은 → 網狀 (reticulate);
- 巢穴 (domatia) → 움 (crypt), 無毛 (glabrous).
- 꽃차례 (Inflorescences, 花序)는
→ 葉腋 (axil)당 1(–2)개이며, 각각 꽃은 하나이고 꽃받침(calyculi)은 두 개. - 아랫쪽 外萼 (Lower calyculus)은
→ 두 개의 작은 엽편(foliar lobes)이 있는 테(rim)로 축소됨. - 위쪽 外萼 (Upper calyculi)은
→ 4개로 갈라짐(four lobed):
→ stipular lobes 2개, 넓은 삼각형;
→ foliar lobes 2개, 작고 좁은 타원형. - 꽃은 → 5장으로 갈라짐(5-merous), 꽃자루가 없음(sessile).
- 꽃부리 (Corolla tube, 花冠 ) → 길이 1mm,
- 꽃잎 (lobes, 裂片)은 → ± 선형, 길이 (5–)6–8mm, 너비 2–3mm, 흰색.
- 암술군 (Gynoecium)은
→ 씨방(ovary) 위에 작은 꿀샘 원반 (밀반, nectary disc); - 꽃받침 테(calyx limb) → 짧고(truncate), 디스크보다 짧으며, 털이 없다.
- 암술대 (Style) → 길이 약 10mm;
- 암술두 (stigma) → 2갈래로 갈라짐(bilobed), 길이 약 2mm.
- 꽃밥 (花葯, Anther) → 길이 약 3mm;
- 꽃밥 필라멘트(anther filament) → 길이 약 2mm, inframedifixed (하부중간부착형).
- 열매 (Fruit) → 9–10mm × 7mm, 빨간색;
꽃자루 (花莖, peduncle, 열매꼭지) → 약 길이 2mm, 털이 없음. - 씨앗 (Seed)은 → 넓은 타원형, 길이 약 5mm, 너비 약 4mm, 두께 약 3mm.
Micromorphology :
- 잎사귀가 얇고, 윗면 표피가 20~30mm, 울타리 모양의 중엽이 20~30mm, 해면 같은 중엽이 45~70mm,
- 아랫면 표피가 10~20mm, 전체 두께가 100~130mm;
- 종자 껍질(‘silver skin’)은 후막(sclereids)이 없는 얇은 막(parenchymatous).
Distribution :
- 서부 카메룬의 바코시 삼림보호구역 토착종.
Habitat and ecology :
- 가파른 경사면의 습한 일차 우림, 바위가 많은 토양.
- 해발 160m.
- 조사한 다른 표본: F. Anthony 17(BR!; flowering); 18(BR!; flowering); 16(BR!; flowering).
- 모두 Ivory Coast의 IRD-커피 컬렉션에서 수집, 생식질 엔트리 넘버 C11,
- 카메룬 남서부 지방, 톰벨 지구, 바코시 삼림보호구역, 뭉고 강의 다리에서 동쪽으로 2km,
N4°44′ E9°35′, 160m에서 수집된 cuttings로부터 키웠음.
Etymology :
- 이 이름은 20세기 마지막 30년 동안 IRD에서 커피 육종 연구와 수집 임무를 관리했던
A. Charrier 교수를 기리기 위해 명명되었다. - 1988년부터 1993년까지 그는 the French Office of Genetic Resources (BRG)를 관리했고,
- 1991년부터는 국립 농업 연구소(the National Institute for Agricultural Research, INRA)에서
'식물 유전학 및 육종' 연구 부서의 책임자를 역임했다.

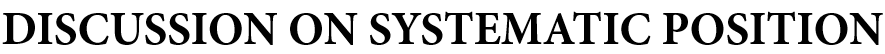
- C. charrieriana의 열매는
→ 두 개의 피렌(pyrenes)(씨 stones)이 있는 육질의 핵과(fleshy drupes)이다. - 각 피렌에는 씨앗이 하나씩 들어 있다.
- 씨앗은
→ 향축으로(adaxially) 평평하고 홈이 있다 (flat and grooved);
→ 배축으로(abaxially) 둥글고 매끄럽다 (rounded and smooth). - 씨앗 껍질(‘silver skin’)은 후막이 없는 얇은 막(parenchymatic without sclereids)이다.
- 이러한 특성의 조합은
두 자매 속인 Coffea와 Psilanthus (씨랜써스)에서 일반적이다. - 꽃 특성을 기준으로 두 속인 Coffea와 Psilanthus는 쉽게 구별할 수 있다.
- Coffea에서는
꽃밥(anthers)과 암술대(style)가 돌출되어 있고,
꽃밥(medifixed)은 축방향으로 고정되어 있으며(submedifixed), 그리고
짧은 필라멘트로 관(tube)과 엽(lobes) 사이의 전환 구역(transition zone)에서
꽃부리(corolla)에 부착되어 있으며,
꽃부리는 반투명하지 않다 (never semi-transparent). - Psilanthus에서는
암술대와 꽃밥은 항상 좁은 꽃부리 통(corolla tube)에 포함되고,
꽃밥은 꽃부리 통(corolla tube)에 고정되어(supramedifixed) 필라멘트 없이 붙어 있으며,
꽃부리 통(corolla tube)은 얇고 종종 반투명하다. - C. charrieriana의 모든 꽃 형태적 특성은 Coffea 속과 일치한다;
따라서 새로운 Coffea 종으로 결론지어진다. - Coffea 속의 특이한 점은
C. charrieriana의 실질 종자 껍질에 후막세포(sclereids)가 없다는 것이다. - 지금까지 조사한 다른 모든 Coffea 종은
후막세포(sclereids)가 있는 실질 종자 껍질(parenchymatic seed coat)을 가지고 있다 (Stoffelen, 1998);
마다가스카르 종에 대한 추가 관찰이 최근에 이루어졌다 (P. Stoffelen, 미공개 데이터). - 그러나 Psilanthus 속에서는
일반적으로 후막세포(sclereids)가 없다;
스클레레이드는 한 종, 즉 P. lebrunianus (R.Germ. & Kesler) J.-F.Leroy ex Bridson에 대해서만
보고되었다 (Stoffelen, 1998). - Coffea charrieriana는 작은 잎, 꽃관, 꽃관 소엽, 꽃밥, 열매, 그리고 몇 개의 2차 맥(secondary nerves)만 있는 얇은 잎으로 인해 다른 모든 중앙 아프리카 Coffea 종과 다르다 (Table 1 참조).
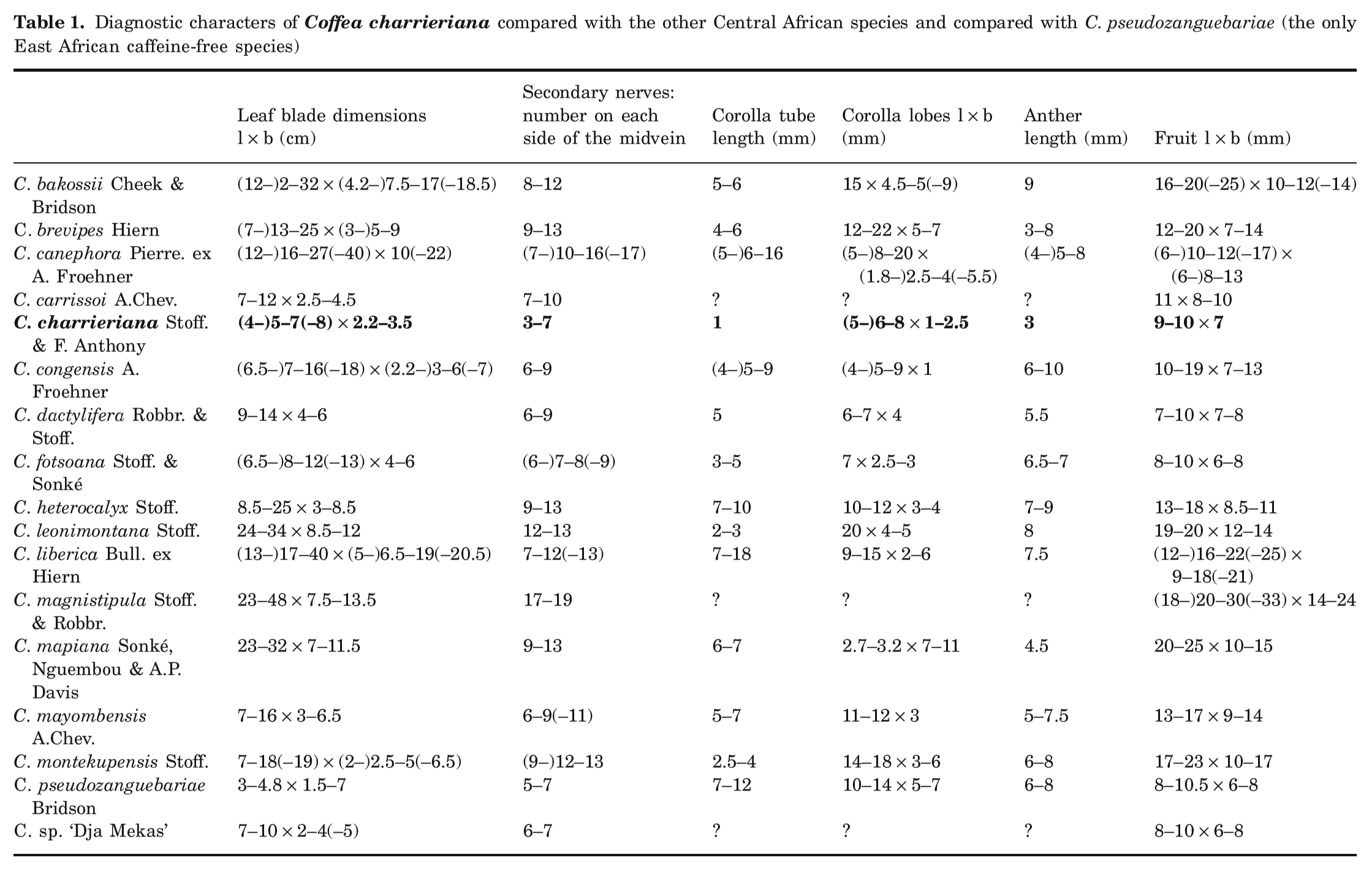
- C. charrieriana의 잎은 매우 얇으며 두께가 100~130mm에 불과하다 (Figure 2).
- 이 점에서
다른 아프리카 Coffea 종 (C. brevipes Hiern: 170~210mm, C. heterocalyx Stoff.: 180~220mm,
C. liberica Bull. ex Hiern: 220~250mm,
C. canephora: 165~230mm) 보다
P. ebracteolatus Hiern (110~136mm)과 더 비슷하다. - 잎사귀의 전반적인 해부학적 구조
(상부 표피(upper epidermis),
책상의 엽육 (palisade mesophyll),
해면 모양의 중엽 (spongy mesophyl),
하부 표피(lower epidermis))는
여기에서 연구한 다른 Coffea 종과 P. ebracteolatus Hiern과 유사하다. - C. charrieriana와 다른 Coffea 종의 주요 차이점은
책상 엽육 (palisade mesophyll), 해면 모양의 엽육 (spongy mesophyl), 표피 세포(epidermis)가
다른 Coffea 종보다 작다는 것이다 (Figure 2 참조).
- Coffea 속 내에서 C. charrieriana는
임시 분류군인 C. sp. ‘Dja Mekas’(Stoffelen, 1998; Stoffelen et al., 1999)와 가장 닮았지만,
털이 많은 가지들 (pubescent branchlets),
얇은 잎사귀들 (thin leaf blades),
더 작은 잎 끝 (smaller leaf acumen),
종자 껍질에 후막세포의 부재 (absence of sclereids in the seed coat)에서 다르다. - C. charrieriana에 대한 추가 관심은
화학 분석 결과 콩에 자연적으로 카페인이 없다는 사실이 밝혀지면서 더욱 커졌다 (Campa et al., 2005). - 이것은 중앙 아프리카에서 알려진 최초의 카페인이 없는 종이기 때문에 매우 주목할 만한 일이다.
- 이전에 카페인이 없는 종은
🌲 마다가스카르의 여러 종 (Bertrand, 1902; Anthony, Clifford & Noirot, 1993)과
🌲 동아프리카의 한 종
C. pseudozanguebariae Bridson
(Hamon, Anthony & Le Pierrès, 1984; Anthony et al., 1993)에서만 알려져 있었다. - 그럼에도 불구하고
C. charrieriana는 짧은 꽃자루와 더 작은 꽃부리와 꽃밥 때문에
C. pseudozanguebariae와 쉽게 구별할 수 있다 (Table 1). - 논의 중인 종의 또 다른 원래 생화학적 특성은
자당 (sucrose) 함량 (3.8% dmb)으로,
Coffea 아속에서 발견되는 가장 낮은 값이다 (Campa et al., 2004). - 새로운 종의 발견은 아프리카 숲의 탐사와 목록이 끝나지 않았다는 것을 다시 한번 보여준다.
- 커피와 같은 경제적으로 중요한 작물에서도 새로운 종이 계속 발견되고 있다.
- 카메룬의 코페아 종의 약 절반이 지난 10년 동안 기술되었다.
- 이는 또한 유전자원의 장기 보존을 위한 보존 프로그램을 시행해야 할 필요성을 지적한다.
REFERENCES
- Anthony F. 1992.
Les ressources génétiques des caféiers: collecte, gestion d’un conservatoire et évaluation de la diversité génétique. Paris: IRD, Série TDM n° 81. - Anthony F, Astorga C, Berthaud J. 1999.
Los recursos genéticos: las bases de una solución genética a los problemas de la caficultura latinoamericana.
In: Bertrand B, Rapidel B, eds. Desafíos de la caficultura centroamericana. San José: IICA/PROMECAFE-CIRAD-IRD-CCCR France, 369–406. - Anthony F, Clifford MN, Noirot M. 1993.
Biochemical diversity in the genus Coffea: chlorogenic acids, caffeine and mozambioside contents. Genetic Resources and Crop Evolution 40: 61–70. - Anthony F, Couturon E, de Namur C. 1985.
Les caféiers sauvages du Cameroun. Résultats d’une mission de prospection effectuée par l’ORSTOM en 1983. In: Proceedings of the 11th International Scientific Colloquium on Coffee. Montpellier: ASIC, 495–505. - Anthony F, Dussert S, Dulloo E. 2007.
The coffee genetic resources. In: Engelmann F, Dulloo E, Astorga C, Dussert S, Anthony F, eds. Complementary strategies for ex situ conservation of Coffea arabica genetic resources. A case study in CATIE, Costa Rica. Rome: Bioversity International, Topical Reviews in Agricultural Biodiversity, 12–22. - Bertrand G. 1902.
Recherche et dosage de la caféine dans plusieurs espèces de café.
Bulletin de la Société de Pharmacie 5: 283–285. - Bridson D, Verdcourt B. 1988.
Coffea. In: Polhill RM, ed. Flora of tropical East Africa (part 2). Rotterdam: A.A. Balkema, 703–723. - Campa C, Ballester JM, Doulbeau S, Dussert S, Hamon S, Noirot M. 2004.
Trigonelline and sucrose diversity in wild Coffea species. Food Chemistry 88: 39–43. - Campa C, Doulbeau S, Dussert S, Hamon S, Noirot M. 2005.
Diversity in bean caffeine content among wild Coffea species: evidence of a discontinuous distribution. Food Chemistry 91: 633–637.
https://doi.org/10.1016/j.foodchem.2004.06.032

- Cheek M, Csiba L, Bridson D. 2002.
A new species of Coffea (Rubiaceae) from western Cameroon. Kew Bulletin 57: 675–680. - Davis AP. 2001.
Two new species of Coffea L. (Rubiaceae) from eastern Madagascar. Kew Bulletin 56: 479–489. - Davis AP, Bridson DM, Rakotonasolo F. 2005.
A reexamination of Coffea subgenus Baracoffea and comments on the morphology and classification of Coffea and Psilanthus (Rubiaceae-Coffeeae).
In: Keating RC, Hollowell VC, Croat T, eds. Festschrift for William G. D’Arcy: the legacy of a taxonomist (monograph in Systematics Botany 104). Missouri: MBG Press, 398–420. - Davis AP, Govaerts R, Bridson DM, Stoffelen P. 2006.
An annotated taxonomic conspectus of the genus Coffea (Rubiaceae).
Botanical Journal of the Linnean Society 152: 465–512. - Davis AP, Mvungi EF. 2004.
Two new endangered species of Coffea (Rubiaceae) from the Eastern Arc Mountains (Tanzania) and notes on associated conservation issues. Botanical Journal of the Linnean Society 146: 237–245. - Davis AP, Rakotonasolo F. 2000.
Three new species of Coffea L. (Rubiaceae) from Madagascar. Kew Bulletin 55: 405–416. - Davis AP, Rakotonasolo F. 2001a.
Two new species of Coffea L. (Rubiaceae) from northern Madagascar. Adansonia 23: 337–345. - Davis AP, Rakotonasolo F. 2001b.
Three new species of Coffea L. (Rubiaceae) from NE Madagascar. Adansonia 23: 137–146. - Davis AP, Rakotonasolo F. 2003.
New species of Coffea L. (Rubiaceae) from Madagascar. Botanical Journal of the Linnean Society 142: 111–118. - Hamon S, Anthony F, Le Pierrès D. 1984.
La variabilité génétique des caféiers spontanés de la section Mozambicoffea A. Chev. 1). Précisions sur deux espèces affines: Coffea pseudozanguebariae Bridson et C. sp. A Bridson. Adansonia 2: 207–223. - Holmgren PK, Holmgren NH, Barnett LC. 1990.
Index Herbariorum. Part I: the herbaria of the world. Regnum Vegetabile 120: 1–693. - Sonké B, Stoffelen P. 2004.
Une nouvelle espèce de Coffea L. (Rubiaceae) du Cameroun, avec quelques notes sur ses affinités avec les espèces voisines. Adansonia 26: 153–160. - Stoffelen P. 1998.
Coffea and Psilanthus (Rubiaceae) in tropical Africa: a systematic and palynological study, including a revision of the West and Central African species. PhD thesis, K.U. Leuven. - Stoffelen P, Cheek M, Bridson D, Robbrecht E. 1997.
A new species of Coffea and notes on mount Kupe. Kew Bulletin 52: 989–964. - Stoffelen P, Robbrecht E, Smets E. 1999.
A new species of Coffea (Rubiaceae) from central Africa, with notes on tentative other taxa. Systematics and Geography of Plants 69: 119–124. - Vega FE, Rosenquist E, Collins W. 2003.
Global project needed to tackle coffee crisis. Nature 425: 343.

'Coffee Botany' 카테고리의 다른 글
| 스테노필라 커피의 플레이버 테이스팅 (9) | 2024.08.30 |
|---|---|
| 스테노필라 커피 (Coffea stenophylla)의 재발견 (3) | 2024.08.29 |
| 코페아 앤서니 (Coffea anthonyi) (2) | 2024.08.27 |
| 리베리카 커피 (2) | 2024.08.15 |
| 커피나무와 유전학 서론 (23) | 2024.08.09 |




댓글