
카메룬(Cameroon)과 콩고 공화국(Republic of Congo)의 코페아 앤서니 Coffea anthonyi Stoff. & F. Anthony가 공식적으로 기술된다. 형태학적, 생화학적 및 분자적 특성, 아이소자임(동질효소, isozyme) 다양성 및 계통발생론적 관계가 논의된다. 이 새로운 이배체 종은 동아프리카 C. eugenioides 및 이질사배체 재배 C. arabica와 강한 관계가 있다. 따라서 C. arabica의 조상 종과 관련이 있을 수 있다. 이 새로운 종은 자가-화합적(self-compatible)이며, 이는 Coffea 속 내에서 희귀한 특성이다. 이 새로운 소엽의 중앙 아프리카 종은 계통발생론적 관점 뿐만 아니라 육종 관점에서도 특히 관심을 끈다.
■ INTRODUCTION
- Coffea는 Rubiaceae (꼭두서니科)에 속하며, 많은 종에서 알 수 있듯이 다양성이 높다.
- 최근에 많은 새로운 분류군이 발견되었다
(Stoffelen & al., 1996, 1997a, b, 1999, 2008; Davis, 2001; Davis & Rakotonasolo, 2000,2001a, b, 2003; Cheek & al., 2002; Davis & Mvungi, 2004; Sonke & Stoffelen, 2004). - 지금까지 103개의 Coffea 종들이 아프리카 대륙과 Madagascar의 숲에서 식별되었다 (Davis & al., 2006).
- 모든 종은 다년생 목본 관목(shrubs) 또는 교목(trees)이며, 그리고 형태, 크기 및 생태적 적응성이 크게 다르다.
- 그들은 이배체(diploid) (2n = 2x = 22) 이며,
C. arabica L.만이 이질사배체(allotetraploid) (2n = 4x = 44)로서 예외적이다
(Krug & Carvalho, 1951; Lashermes & al., 1999). - 상업적 커피 생산은 주로 두 가지 종들
C. arabica 및 C. canephora Pierre ex A. Froehner에 의존하며,
그 중 C. arabica에 더 나은 품질의 커피가 관련되어 있다. - 이배체 종으로부터의 유전자들이 종간교잡화 (interspecific hybridization)에 의해
C. canephora 또는 C. arabica 게놈으로 이전될 수 있다. - 그러나, 커피 나무들의 천연 개체군들은 수십년 전부터 아프리카와 마다카스가르에서 발생하는
대량 삼림벌채에 의해 위협 받아 오고 있다. - 기존 유전자은행들에 보존되는 다양성의 부족 때문에,
국제 기구들 (FAO and IPGRI)과 프랑스 조직들 (IRD ex-ORSTOM, CIRAD, MNHN)은
커피 생식질을 수집하는 협력적 노력을 해왔다. - 지난 40년 동안, IRD는
Ethiopia (Guillaumet & Halle, 1978),
Kenya (Berthaud & al, 1980),
Tanzania (Anthony & al., 1987),
Central African Republic (Berthaud & Guillaumet, 1978),
Cameroon (Anthony & al., 1985),
Republic of the Congo (Namur & al, 1987),
Ivory Coast (Berthaud, 1986) and Guinea (Le Pierres & al, 1989)에서
컬렉션 미션들을 조직하고 수행했다. - 약 20가지의 분류군에 속하는
총 7,800개의 야생 커피 유전자형들이 수집되었고,
Ivory Coast의 필드 유전자은행에 도입되었다 (Anthony, 1992). - 여기서, 우리는 새로운 커피 종 C. anthonyi를 기술한다.
- 이는
Cameroon 남부지방 (Anthony & al, 1985)과
카메룬과 콩고공화국 (Namur & al, 1987) 사이의 국경지대에서
발견되었다. - C. anthonyi의 발견은 중앙 아프리카 서부에 중요한 커피 다양성 센터의 존재를 확인해준다.
- 최근 몇 가지 새로운 분류군이 기술되기 전까지
카메룬에서 발견되는 커피 종의 수는 과소평가되었다 (Stoffelen & al. 1996,1997a, b, 1999,2008; Cheek & al. 2002; Sonke & Stoffelen 2004; Sonke & al. 2006). - 1995년에, 단지 5개 종들만이 알려졌다; 이 숫자는 이제 14종으로 증가되었다.
- C. anthonyi의 동질효소 다양성(Isozyme diversity)을 평가하고,
C. brevipes Hiera, C. canephora, C. liberica Hiera 및 C. mayombensis A. Chev와 같은
잘 알려진 중앙 아프리카 종의 아이소자임 다양성과 비교한다. - 나아가 그 새로운 종에 대한 생화학적, 유전적 및 분자적 데이터를 검토한다.
- C. anthonyi의 식물 재료가 주로 몽펠리에의 IRD에서 커피 연구자들에 의해 수년간 재배되고 연구되었기 때문에 많은 출판된 데이터가 존재한다.
- 지금까지 이러한 데이터는 공식적인 분류명을 사용하여 발표되었으므로 그 수용되는 이름과 연결될 필요가 있다.
■ MATERIALS AND METHODS
Germplasm collection. ─
- 생식질 컬렉션 미션은
Cameroon에서 1983년 1월-3월 (Anthony & al, 1985) 그리고
Republic of the Congo에서 1985년 8월-9월에(Namur & al, 1987) 이뤄졌다. - 그 새로운 종은
Cameroon의 세 군데 (C27, C29, C30), 그리고
Republic of the Congo의 두 군데(Cg15, Cg17)에서 발견되었다 (Fig. 2). - 수집된 재료는
C27에서 5개의 cuttings,
C29에서 1개의 cutting,
C30에서 3개, Cg15에서 2개, 그리고
Cg17에서 48개였다. - 씨앗들도
C27 (한 나무에서 2개 종자), Cg15 (한 나무에서 4개 종자), 그리고 Cg17 (4그루 나무로부터 17개 종자)에서
수집되었다. - 그 개체군에 관한 지역 데이터는 다음과 같다 :
C27 (Moloundou-1 population): N2°03′ E15°05′;
C29 (Moloundou-2 population): N2°04′ E15°00′;
C30 (Moloundou-3 population): N2°06′ E14°56′;
Cg15 (Souanké-Gabon-1 population): N2°07′ E14°00′;
Cg17 (Souanké population): N2°03′ E14°08′.
Study of herbarium material. ─
- 6개의 식물표본집 표본들이 식물학적 기술을 위해 연구되었다 (BR, BRLU, K, LISC, P, WAG).
- 이 식물표본집들로부터 모든 다른 Coffea의 자료들이 그 새로운 종을 위해 검토되었다.
Isozyme analysis. ─
- 연구된 자료는
31개의 C. anthonyi 액세션들과
35개의 C. mayombensis 액세션들로부터의 어린 잎들로 이뤄졌는데,
각 종은 두 개체군들이 대표하였다 (Table 1) (Anthony & al, 1985; Namur & al, 1987). - 전기영동(Electrophoresis) 데이터가 Berthaud (1986)에 의해 설명된 방법들을 따라 기록되었다.
- 효소들은 다음과 같았다.
α-esterase (EST),
isocitrate dehydrogenase (ICD),
phosphoglucoisomerase (PGI),
phosphoglucomutase (PGM), 그리고
6-phosphogluconate dehydrogenase (PGD). - 총 33개의 대립유전자 빈도(allelic frequencies)가 각 샘플마다 평가되었다.
- 그런 다음 개체군별 빈도들이 계산되었고
Statistica 소프프웨어 (© Statsoft, Inc.)의 joining method를 사용하여 분석되어
계층수(hierarchical tree)를 생성하였다. - 중앙아프리카 토종인 3개의 잘 알려진 종들 (즉, C brevipes, C canephora, C. liberica)에 관한 데이터는
Berthaud (1986)로부터 가져왔고, 비교분석에 포함되었다 (Table 1).

Controlled self-pollination. ─
- 통제된 교잡화(controlled hybridisations) 방법이
종간 하이브리드를 사용하는 커피 육종 프로그램들에서(Van der Vossen, 1985),
특히 Ivory Coast에서 흔히 사용된다 (Louarn, 1992). - 자가수분(Self-pollination)은
개화(anthesis)와 수분(pollination) 전에 꽃 피는 가지들을 봉지로 씌워서(by bagging) 통제할 수 있다. - 개화 시, 자가수분을 유도하기 위해 가지들을 흔든다.
- 봉지들은 이틀 후에 제거한다.

■ DESCRIPTION OF THE NEW SPECIES
Coffea anthonyi Stoff. & F. Anthony, sp. nov. ─
- Holotype: Anthony F. 20 (BR0000006920555),
grown at Divo coffee genebank, Ivory Coast,
from cuttings collected
in the Republic of the Congo, at Souanke population, 1999 (Fig. 1). - Diagnosis: Coffea anthonyi differt ab specie
Africae occidentalis et Centralis prope foliis minutis et obovatis.
(잎이 작고 난형이라는 점에서 서부 및 중앙아프리카의 종과 다르다.) - Coffea sp. 'Moloundou', informal name in
Anthony, 1992: 194;
Anthony & al., 1993: 65-70;
Cros & al, 1995: 15-18, 1998: 110-115;
Cros, 1996: 14-138;
Lashermes & al, 1996: 628-630, 1997: 948-954, 1999: 260-164;
Stoffelen, 1998: 125;
Combes & al, 2000: 1179;
Noirot & al, 2003: 710-713;
Campa & al, 2004: 40-43, 2005a: 634-636,2005b: 135-139;
Poncet & al, 2004:1072-1078, 2005: 2-5;
Anthony & Lashermes, 2005: 210-214. - Coffea sp. 'Dja Mekas', informal name in
Stoffelen, 1998: 123-124;
Sonke & Stoffelen, 2004: 157.
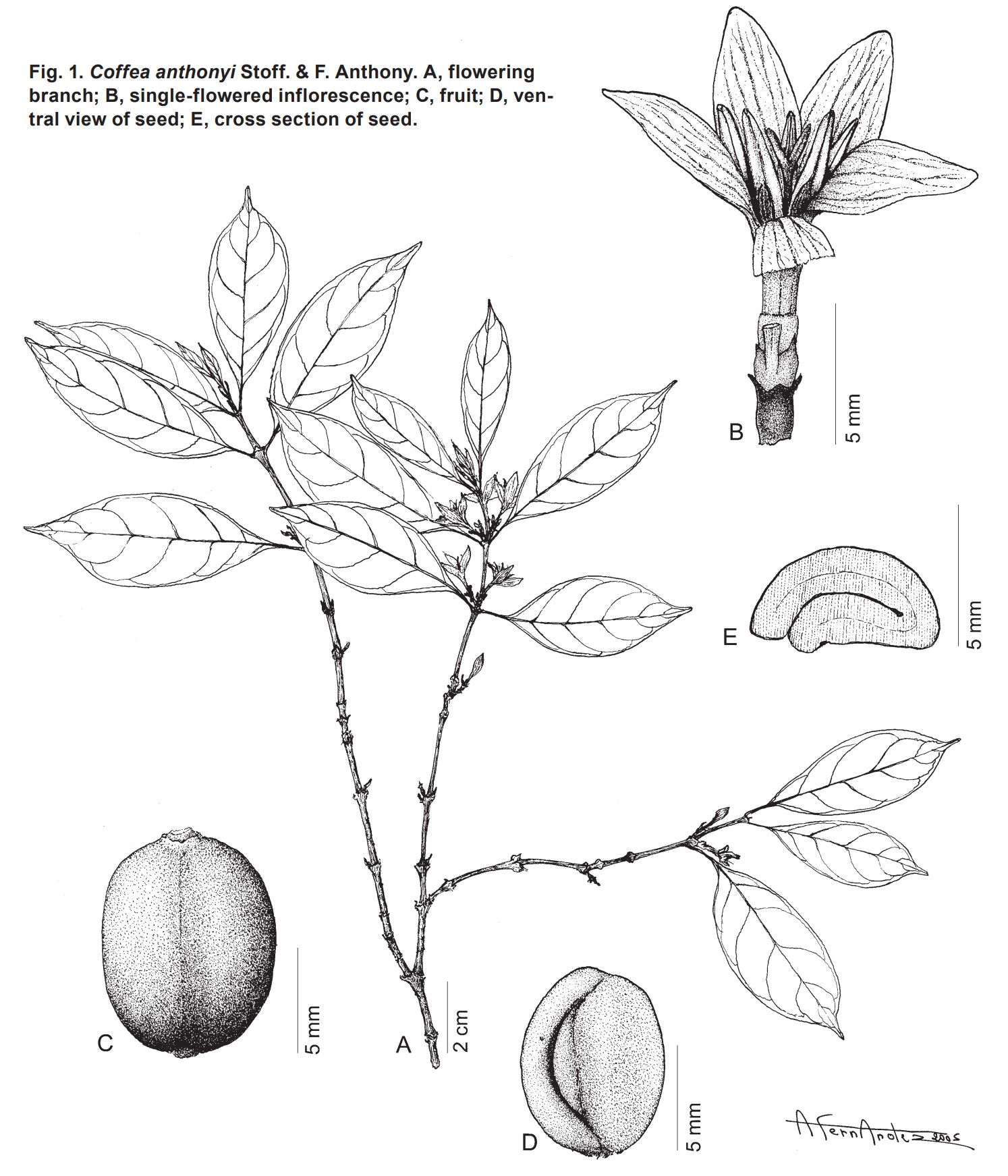
- 키가 최대 2m인 작은 관목(Small shrub), 어린 가지는 털이 없다(glabrous).
- 잎사귀(Leaf-blades)는 도란형(obovate)이고 드물게 타원형이며, 길이 5-9(-10)cm, 너비 2.2-4(-5)cm이다;
- 잎 밑부분(base)은 쐐기 모양(cuneate);
잎 끝(tip)이 뾰족하고(acuminate), 뾰족한 끝(acumen)은 길이 5-8(-10)mm이며, 끝이 둥글다(rounded tip);
잎의 기부 절반에 있는, 주맥(midrib)과 2차 맥의 접합부에 위치한 작은 crypt domatia(巢穴)는 작은 구멍이 있는 능선(ridges with minute openings)으로 보이며, 털이 없다;
주맥 양쪽에 5-7개의 2차 맥이 있다. - 잎 꼭지(Petioles)는 길이 5-10mm이다.
- 턱잎(Stipules)은 삼각 모양이고(deltate), 가지(limb) 길이는 약 2mm이며, 섬모가 있다(ciliate).
- 꽃차례(Inflorescences)는
짧은 꽃자루가 있고(pedunculate), 겨드랑이(axil)당 1-2개이며,
각각 1-2개의 꽃과 2-3개의 꽃받침(calyculi)이 있다; - 꽃자루는 작다(peduncle minute); 꽃받침은 4개로 갈라지고, 2개의 턱잎 갈래와 2개의 작은 foliar lobes가 있다.
- 꽃은 5 갈래이다(5-merous).
- 꽃받침(花萼, Calyx)은 통 모양이다(truncate).
- 꽃부리(화관, Corolla)은 흰색이다; 관 길이는 약 5mm이다; 엽은 길이 6-7mm, 너비는 약 3mm이다.
- 수술(Stamens)은 꽃밥이 있고(with anthers) 길이 4.5-5mm, 필라멘트(filaments) 길이는 약 0.5mm이다.
- 암술군(Gynoecium)은 털이 없다(glabrous). 작은 고리 모양의 원반(Small annular disc)이 있다.
- 암술대(花柱, Style)는 길이 약 10mm이다; 암술머리(stigma)는 2개로 갈라지고, 길이는 약 2mm이다.
- 열매차례(Infructescences)는 꽃자루가 있고(with peduncle) 길이 2.5-3mm이다.
- 열매는 빨간색이고 길이 9-10mm, 너비 6-8mm이다.
- 씨앗은 넓은 타원형이고 길이 7-8mm, 너비 5.5-6mm, 두께 약 3mm이다.
- 실버스킨은 유세포 조직(parenchymatous)이며 평행하게 모인 길쭉한 후막세포(sclereids)가 있다;
- 스클레레이드에는 원형 또는 렌즈형 구멍들(perforations)이 있다.




Material studied. —
- Cameroon:
Breteler F.J. 853 (K, LISC, WAG), 5 km east of Bertoua, Batouri-Betaré-Oya, N4°35′ E13°41′;
Breteler F.J. 2189 (LISC, P, WAG), Gounté, 27 km on road Bertoua-Bétaré-Oya, N4°42′ E13°49′;
Sonké B. 1412 (BR, BRLU): réserve de faune du Dja Mékas, N3°11′ E12°33′. - Republic of the Congo:
Anthony F. 19 (BR), grown in Divo coffee genebank, Ivory Coast from cuttings
collected in the Republic of the Congo, at Souanké-Gabon-1 population, N2°07′ E14°00′;
Anthony F. 20 (BR), grown in Divo coffee genebank, Ivory Coast from cuttings
collected in the Republic of the Congo, at Souanké population, N2°03′ E14°08′;
Anthony F. 21 (BR), grown in Divo coffee genebank, Ivory Coast from cuttings
collected in the Republic of the Congo, at Souanké population, N2°03′ E14°08′.
Distribution and habitat. —
- Coffea anthonyi는 카메룬 남동부 및 콩고공화국 북서부의 토착종이다 (Fig. 2).
- 거의 모든 자료들이, 이 종은 “1차” 우림에서 발견된다고 한다;
Breteler 853은
“gallery forest, mostly secondary, marshy soil (갤러리 숲, 대개 2차적인, 습지성 토양)”이라고 한다. - 야생에서, C. anthonyi는 종종 다른 Coffea 또는 Psilanthus 종들과 연관된다.
- C. canephora (population Cg15),
C. liberica var. dewevrei (De Wild. & T. Durand) Lebrun 및
Psilanthus mannii Hook. f. (population C27)와 함께, 그리고
P. mannii (populations C29, C30 and Cg17)와 함께 발견된 바 있다. - Cg17 개체군은 Triplochiton scleroxylon K. Schum (오베체)의 오래된 숲으로 덮인 경사면에서 약 1,000m² 면적에서 발견되었으며, 바위가 많은 협곡과 수원이 있는 바위가 많은 토양에서 발견되었다.
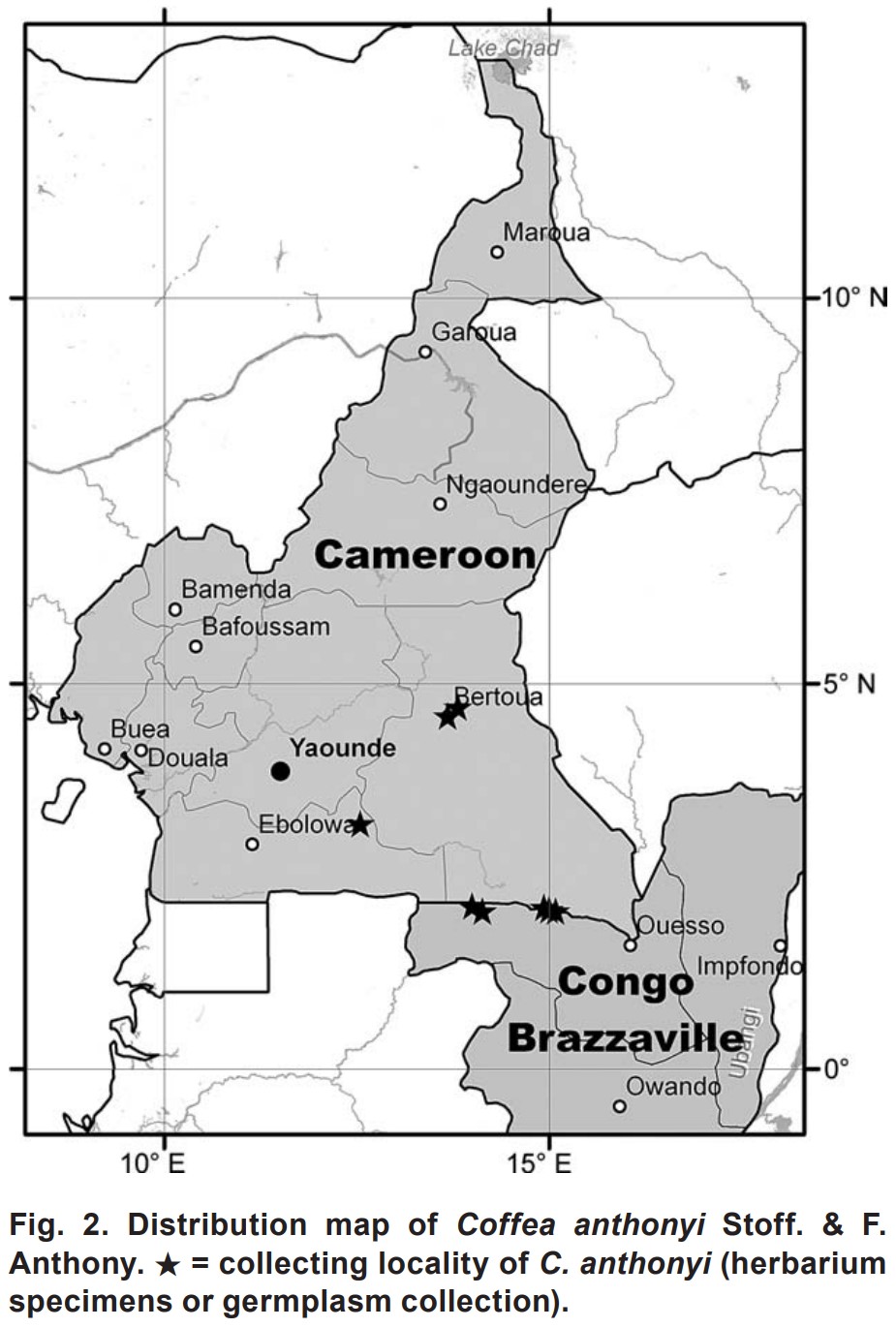
History of species recognition. —
- 이 새로운 종은 잎과 열매의 형태가 기술된 어떤 커피 종과도 맞지 않아
IRD 수집가에 의해 Coffea sp. nov.로 식별되었다 (Lebrun, 1941; Chevalier, 1947; Bridson & Verdcourt, 1988). - 이전 출판물에서 이 종은 Coffea sp. ‘Moloundou’로 명명되었다.
- A. Charrier (WAG의 Breteler 2189 & 853 폴더에 있는 서면 메모)에 의해
나중에 비공식 분류군 Coffea sp. ‘Dja Mekas’ (Stoffelen, 1998; Stoffelen & al, 1999)로 명명된
식물표본 자료와 연결되었다. - Coffea sp. ‘Dja Mekas’는 공식적으로 기술되지 않았는데, 열매를 맺는 표본만 이용가능했고, 어떤 경우에는 열매를 맺는 Coffea와 Psilanthus Juss. ex M. Roem. 개체들이 구별하기 매우 어렵기에 꽃을 관찰하는 것이 Coffea 종을 식별하는데 절대적으로 필요하기 때문이다.
- 1999년 카메룬과 콩고 공화국에서 수집하여 코트디부아르의 Divo 커피 컬렉션에서 재배한 새로운 종의 식물표본 자료가 몽펠리에의 IRD로 배송되었다.
- 이 새로운 자료에 대한 연구를 통해,
해당 種이 과학에 새로운 종이며,
Coffea sp. ‘Moloundou’와 Coffea sp. ‘Dja Mekas’가 '동의어'라는 것이 확인되었다. - The Democratic Republic of the Congo의 한 컬렉션 (Sapin B2 [BR], Katola, Sandoa)은 이 새로운 종을 연상시키지만, 재료가 너무 부족하여 그 정체에 관해 확신하기 어렵다 (immature fruits).
Comparison with morphologically similar Coffea species. —
- 새로운 종은 중앙 아프리카에서 온 몇 안 되는 작은 잎을 가진 커피 종 중 하나이다.
- 중앙 및 서부 아프리카에서 두 가지 다른 작은 잎을 가진 종만이 기술되었다:
C. kapakata (A. Chev.) Bridson (Chevalier, 1947; Bridson, 1994)과
C. charrieriana Stoff. & F. Anthony (Stoffelen & al, 2008). - Coffea kapakata는 C. anthonyi와 구별할 수 있는데,
전자는 전형적인 타원형 잎, 엽상 꽃받침 및 늑골 모양의 열매가 있는 앙골라의 사바나 (savannah) 종이기 때문이다. - Coffea charrieriana는
타원형 또는 넓은 타원형, 드물게 도란형 잎과
매우 작은 꽃밥(약 3mm 길이)을 가지고 있는 반면, - C. anthonyi는 도란형, 드물게 타원형 잎과 4.5-5mm 길이의 꽃밥을 가지고 있다.
- 새로운 종의 형태는
동아프리카 종인 C. eugenioides S. Moore의 형태와 비슷하지만,
🌲 도란형 잎 (C. eugenioides는 타원형),
🌲 짧은 꽃자루 (C. eugenioides는 5-16mm인데, 2-3mm에 불과한) 및
🌲 짧은 꽃밥 필라멘트 (C. eugenioides는 0.8-3mm인데 비해, 0.5mm)로 후자와 다르다.
Isozyme diversity and differentiation. ─
- 중앙 아프리카의 5종에 속하는 237개 커피 액세션들에서
33개 대립유전자의 존재 또는 부재가 기록되었고 (Table 1 참조), 그런 다음 대립유전자 빈도가 개체군별로 계산되었다. - 그 결과 계층적 트리는 여러 수준의 클러스터링을 명확하게 보여준다 (Fig. 3).
- 예상대로 같은 종에 속하는 개체군은 클러스터링의 가장 낮은 수준에서 함께 연관된다 :
C. mayombensis, C. canephora 및 C. anthonyi. - C. brevipes 개체군은 그 다음 수준에서 C. canephora 개체군과 그룹화된다.
- 이 그룹화는 C. brevipes가 C. canephora의 생태형으로 간주될 수 있기 때문에 예상되었다 (Chevalier, 1947; Anthony, 1992; Stoffelen, 1998).
- 클러스터링의 세 번째 수준은 C. mayombensis 개체군을 C. canephora-C. brevipes 그룹과 연관시킨다.
- 네 번째 수준에서 C. liberica는 이 다중특이적 그룹과 연관되는 반면,
마지막 수준에서 C. anthonyi는 다른 종과 그룹화된다. - 이 분류는 분자 데이터(N'Diaye & al, 2005)를 기반으로 추정된
C. canephora와 C. liberica 사이의 높은 유전적 분화(0.74)를 확인한다. - C. anthonyi는 계층적 트리의 가장 높은 수준에서만 다른 종과 그룹화되므로 같은 지역에서 유래한 다른 종과의 차이는 C. canephora와 C. liberica 사이의 유전적 차이보다 훨씬 더 높을 수 있다.
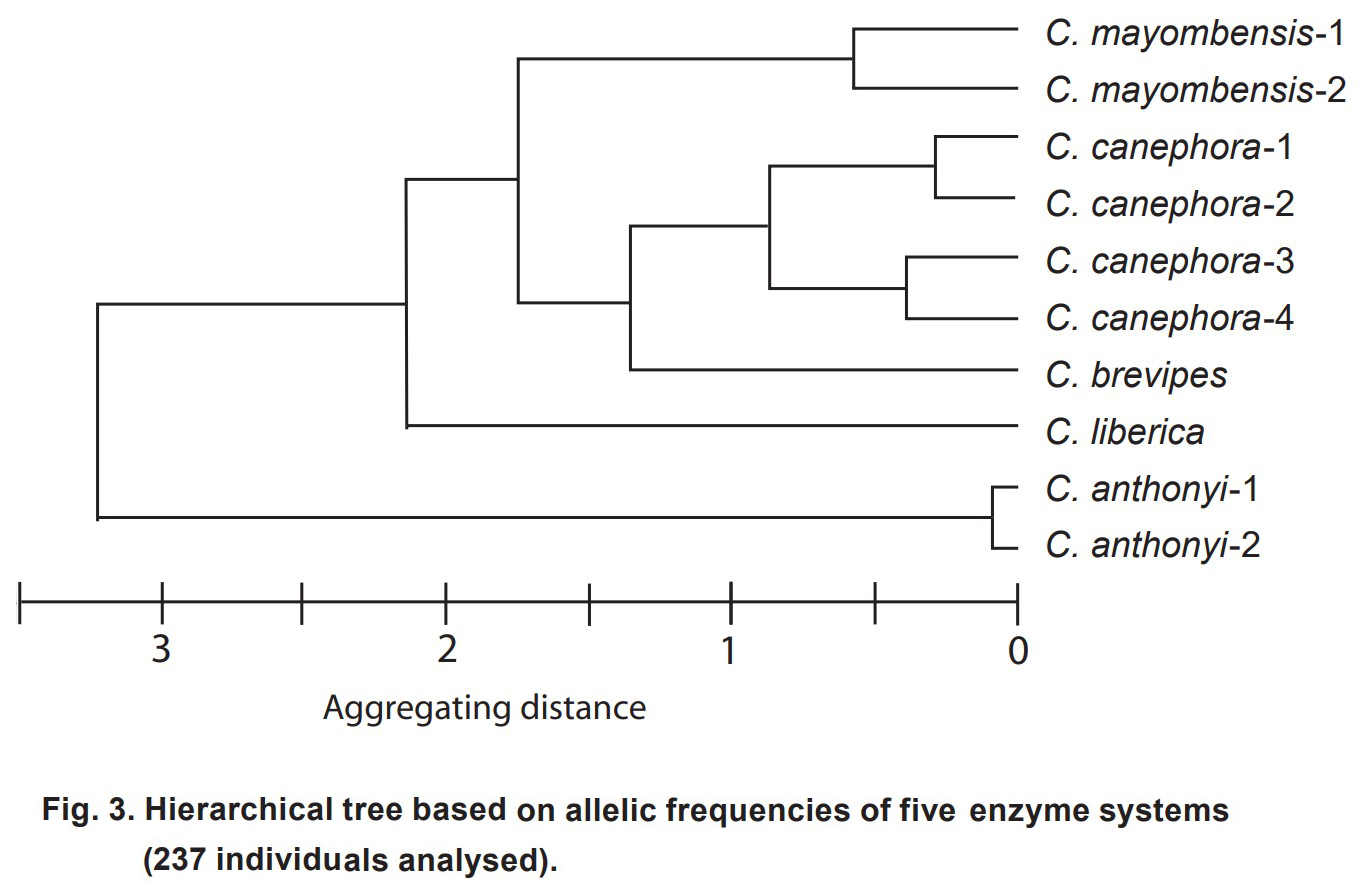
- C. anthonyi의 이러한 독특한 위치는
다른 종의 개체군에서는 발견되지 않은 두 가지 특정 대립유전자(즉, PGD1-X, ICD-I)에 의해 뒷받침된다. - 4가지 대립유전자 (즉, EST-H, PGD2-G, PGI-D, PGM-F)는
C. anthonyi 개체군에서 고정되었고
다른 개체군에서는 다양한 빈도로 발견되었다. - 2가지 대립유전자 (즉, EST-F, PGM-E)는
C. anthonyi의 1~2개 액세션에서 발견되었으며,
이는 이형접합 유전자형 (heterozygous genotypes)에 해당한다. - 6가지 고정 대립유전자(이중 2가지는 커피에서 발견된 적이 없는 것들)의 존재는
그 새로운 종의 오리지날 효소 패턴을 가리켰다. - 연구에서 확인된 다른 대립유전자들(25)은
다른 종의 적어도 1개 액세션에는 존재했지만, C. anthonyi에는 존재하지 않았다. - 또 다른 연구에서는 the acid Phosphatase system에서
특정 패턴(Z4)도 발견되었다 (Berthaud, 1986).
Biology: self-compatibility, ploidy and phenology. ─
- 이 새로운 종의 자가화합성(self-compatibility)은 C27 개체군의 방문 중에 의심되었다.
여기에서 과일 하나를 맺은 단일 커피나무(높이 2m)와 두 개의 어린 나무(높이 20-40cm)가 발견되었다.
넓은 지역에 걸쳐 집중적인 수색을 수행했지만 다른 성체 나무는 발견되지 않았다. - 생식 식물이 이처럼 희소한 것은 커피 개체군에서 매우 드문데,
이배체 종의 자가불화합성(self-incompatibility)은 수정을 위해 두 그루의 나무가 있어야 하기 때문이다. - 게다가 단일 과일에서 씨앗 두 개를 수확한 것은 두 개의 난자가 성공적으로 수정되었음을 나타낸다.
- 다시 말하지만, 곤충에 의한 무작위 수분 때문에 자가화합성 커피 종에서는 이런 일이 흔하지 않다.
- 그런 다음 자가화합성(Self-compatibility)은
Ivory Coast의 야외 유전자 은행에서 통제된 자가수분(self-pollination) 실험을 통해 확인되었고(Anthony, 1992),
열매 세트(29.7%)는 자가화합성 종인 C. arabica(Berthaud, 1978)의 오픈-수정(open-pollinations)의 열매 세트와 유사했다. - 번식 주기(reproduction cycle)는 약 8개월이 걸릴 것으로 추정되었다.
- C. anthonyi의 자가화합성(self-compatibility)의 결과로 이형접합성(heterozygosity)은 낮다.
- 세 가지 액세션에서 보이는 이형접합성 패턴은 두 가지 뿐이었다.
- 이는 공우성 분자 마커(co-dominant molecular markers)를 기반으로 자가화합성 및 자가불화합성 종에서 이형접합성을 비교한 이전 결과를 확인한다.
- RFLP (Restriction Fragment Length Polymorphism) 마커에 의해 검출된 이형접합성 유전자좌의 비율은
C. anthonyi에서 4%였지만
C. eugenioides에서는 23%,
C. congensis A. Froehner에서는 21%,
C. canephora에서는 36%였다 (Lashermes & al, 1999). - 이 비율은 SSR (Simple Sequence Repeat) 마커를 사용하면 더 높았다 (10%)(Poncet & al, 2004).
- 자가화합성은 지금까지 다른 두 커피 종에서 보고되었다 :
이질사배체(allotetraploid) 종인 C. arabica (Carvalho & al, 1991)와
카메룬의 이배체(diploid) 종인 C. heterocalyx Stoff (Coulibaly & al, 2002)이다. - 자가화합성 식물에서 생산성은
불화합성 식물보다 환경 요인에 덜 의존하는데,
이는 최소 두 개의 교차-화합성 (cross-compatible) 식물의 현장 연합,
동시적 개화 (synchronous flowering),
바람과 곤충과 같은 수분 매개체의 가용성에 의해 수정 성공 (fecundation success)이 덜 영향을 받거나 전혀 영향을 받지 않기 때문이다. - C. anthonyi 또는 C. heterocalyx로부터
C. canephora 재배품종으로 자가화합성을 이입하면 동형접합 재배품종 (homozygous cultivars)의 선발과 종자에 의한 그들의 분산이 용이해질 것이다. - C. anthonyi의 배수성(ploidy)에 대한 정보는
공우성 마커(co-dominant markers)를 기반으로 한 분자 연구에서 추론할 수 있다. - 좌위당 최대 두 개의 대립유전자는 항상 동질효소 (isozyme), 미소부수체 (microsatellite) (Combes 등, 2000; Poncet 등, 2004) 그리고 RFLP (Lashermes 등, 1999) 마커를 사용하여 관찰되었다.
- 이는 C. arabica를 제외한 다른 모든 Coffea 종과 마찬가지로
C. anthonyi가 이배체(diploid)라는 가설을 강력히 뒷받침한다. - C. anthonyi의 유전체적 특성(게놈 특성)은 다른 커피 종의 유전체적 특성과 크게 다르지 않다.
- 핵 DNA 함량이 외부(Cros & al, 1995) 또는 내부 표준화(Noirot & al, 2003)를 이용한 유세포 분석(flow cytometry)을 사용하여 추정되었다.
- 내부 표준화를 사용하여, C. anthonyi의 게놈 크기는 약 1.44pg로
Moloundou와 Souanke 개체군 간에 유의미한 차이가 없음을 보여주었다. - 대조적으로 종 간 변이(between species variation)는 존재하고,
C. anthonyi는 C. eugenioides (1.36 pg) 및 C. heterocalyx (1.74 pg)와
다른 것으로 밝혀졌다 (Noirot & al, 2003). - SSR 또는 AFLP (Amplified Fragment Length Polymorphism) 마커를 사용하여,
새로운 종은 C. arabica, C. canephora 및 C. liberica (Combes & al, 2000; Poncet & al, 2004, 2005)와 같은
잘 알려진 종과 유사한 증폭 산물을 보였다. - 여러 종이 공유하는 이러한 마커는
커피의 합의 유전체 지도(consensus genetic map)를 구성하는 데 사용할 수 있을 것이다. - 관련 종 간의 유전자 흐름은 종종 생식 장벽 (reproductive barriers)에 의해 제한된다.
- Coffea에서 가장 흔한 생식 장벽은 감수 분열 동안 염색체 짝짓기 제한 (chromosome pairing limitation)으로 인해 발생한다 (Louarn, 1992). 이로 인해 꽃가루의 생식력(pollen fertility)이 감소한다.
- 커피 나무에서 꽃의 성숙은 비에 의해 유발되며, 개화는 비가 내린 후 5~8일 후에 발생하여 단 하루만 지속된다.
- 비에서 개화까지의 간격 길이가 다르면 종 간의 수분이 제한되고 중요한 생식 장벽이 된다.
- C. anthonyi에서 지연은 7일이다.
- 이는 C. canephora의 경우에도 마찬가지인 반면,
C. eugenioides와 C. arabica의 경우 8일이다 (Baranski, 미공개 자료). - 유전자 흐름에 영향을 미치지 않지만 새로운 종은
C. eugenioides에 비해 두 가지 다른 생물계절학적 차이점 (phenological differences)을 특징으로 한다. - 실제로 과실 성장은 개화 5주 후에 시작되고,
과실은 C. anthonyi의 경우 27주 후에 성숙하는 반면,
C. eugenioides의 경우 이러한 기간은 각각 7주와 23주이다 (Baranski, 미공개 자료).
Biochemical characterisation of the coffee beans. ─
- 이 새로운 종은 콩의 생화학적 조성에서 다른 Coffea 종과 다르지 않다.
- 콩의 카페인 함량은 약 0.6% dmb (dry matter basis)으로 낮으며 (Anthony & al, 1993; Campa & al, 2005a),
C. eugenioides와 다르지 않다. - 클로로겐산(chlorogenic acid) 함량(5.5% dmb)은
Coffea 아속의 두 극단인
C. charrieriana (0.8% dmb)와
Coffea sp. N’koumbala (11.9% dmb)의 중간 수준이다 (Campa & al, 2005b). - 트리고넬린(trigonelline) 함량은
1.6% dmb로 아속 중 가장 높은 수준 중 하나이지만,
C. heterocalyx (1.5% dmb)와 C. eugenioides (1.3% dmb)와 다르지 않다 (Campa & al, 2004). - 마지막으로, 자당 (sucrose) 함량은 5.7% dmb로,
C. heterocalyx (6.2% dmb)와 통계적으로 유사하지만
C. eugenioides (1.1% dmb)보다 상당히 낮다 (Campa & al, 2004).
Phylogenetic relationships. ─
- 커피 종의 계통발생학적 관계가 엽록체(chloroplast)와 리보솜 유전체(ribosomal genomes)의 보존 영역(conserved regions)에서 얻은 데이터를 사용하여 연구되었다.
- cpDNA의 변이는
전체 cpDNA 그리고 atpB-rbcL 유전자 간 영역에 대한 RFLP 분석 (Lashermes & al, 1996)과
trnL-trnF intergenic spacer를 시퀀싱하여 평가했다 (Cros, 1996; Cros & al, 1998). - 다른 데이터는 rDNA의 the internal transcribed spacer (ITS 2) 시퀀스 분석을 통해 얻었다 (Lashermes & al, 1997).
- 이 모든 연구에서 새로운 종은 C. eugenioides와 함께 그룹화되었다.
- C. anthonyi와 C. eugenioides 사이의 rDNA nucleotide-sequence divergence는
같은 종의 개체군 간의 차이(divergence)와 유사하다고 추정되었다 (Lashermes & al, 1997).
두 종의 cpDNA 시퀀스에서 차이가 검출되지 않았다 (Lashermes & al, 1996; Cros & al, 1998). - 나아가, cpDNA polymorphism data는
C. anthonyi와 C. eugenioides를 이질사배체 종 C. arabica와 함께 분류했다 (Cros & al, 1998). - C. arabica, C. anthonyi, 그리고 C. eugenioides 간의 관계는
C. arabica와 여러 잠재적 이배체 조상종들에서 다양한 single-copy probes에 의해 검출된 RFLP 유전자좌의 대립 유전자 구성을 분석하여 확인되었다 (Lashermes & al, 1999; Maurin & al, 2007). - cpDNA는 커피에서 엄격한 모계 유전(maternal inheritance)을 가지므로 (Lashermes & al, 1996),
C. anthonyi 및 C. eugenioides는 이질사배체종 C. arabica의 모계 조상 종과 관련이 있을 수 있다. - C. anthonyi 및 C. eugenioides의 유전적 친밀성(genetic closeness)은
두 종이 분리된 분포 에리어를 가지고 있고 다른 서식지에 적응했기 때문에 자세히 조사해야 한다. - Coffea anthonyi는 카메룬과 콩고 공화국의 저지대 숲(360~650m)에서 서식하는 반면,
C. eugenioides는 콩고 민주공화국 동부, 우간다, 케냐, 탄자니아의 산지림(montane forest)과 산기슭 숲(1,050~2,100m)에서 서식하는 것으로 보고되었다 (Bridson & Verdcourt, 1988; Stoffelen, 1998).
■ CONCLUSION
- 코페아 앤서니 Coffea anthonyi는
형태학적 또는 동질효소 데이터를 사용하여 중앙 아프리카에서 유래한 다른 커피 종과 쉽게 구별할 수 있다. - 그것의 자가-화합성은 Coffea 종에서 드문 특성이며, 유전 구조의 동질화를 촉진했다.
- 결과적으로 C. anthonyi의 이형접합성(heterozygosity)은 자가-불화합성 종보다 상당히 낮다.
- 카메룬과 콩고 공화국의 국경 지역에서 C. anthonyi가 발견된 것은
서부 중앙 아프리카에 커피의 중요한 다양성 센터가 존재함을 확인시켜준다. - 형태학적 및 매크로분자 데이터는
C. anthonyi가 이질사배체 종 C. arabica 및 이배체 동아프리카 종 C. eugenioides와 밀접한 관련이 있음을 분명히 보여준다. - 따라서 C. anthonyi는 C. arabica의 기원을 이해하는 데 중요한 종이 될 수 있다.
■ LITERATURE CITED
- Anthony, F. 1992.
Les ressources genetiques des cafeiers: col lecte, gestion d'un conservatoire et evaluation
de la diver site genetique. Travaux et Documents Microedites, No. 81. ORSTOM, Paris. - Anthony, F., Berthaud, J, Guillaumet, J.L. & Lourd, M. 1987.
Collecting wild Coffea species in Kenya and Tanzania. PI. Genet. Resources Newslett. 69: 23-29. - Anthony, F, Clifford, M.N. & Noirot, M. 1993.
Biochemical diversity in the genus Coffea: chlorogenic acids, caffeine and mozambioside contents.
Genet. Resources Crop Evol. 40: 61-70. - Anthony, F, Couturon, E. & Namur, C. de. 1985.
Resultats d'une mission de prospection effectuee par l'ORSTOM en 1983. Pp. 495-505
in: Proceedings of 11th International Scientific Colloquium on Coffee. Association Scientifique Internationale du Cafe, Lausanne. - Anthony, F. & Lashermes, P. 2005.
The coffee (Coffea ara bica L.) genome: diversity and evolution. Pp. 207-228
in: Sharma, A.K. & Sharma, A. (eds.), Plant Genome: Biodiversity and Evolution, vol. IB, Phanerogams. Science Publishers Inc., New Hampshire. - Berthaud, J. 1978.
L'hybridation interspecifique entre Coffea arabica L. et Coffea canephora Pierre. Obtention et com paraison des hybrides triploi'des, Arabusta et hexaploi'des. Cafe, Cacao, The 22: 3-12. - Berthaud, J. 1986.
Les ressources genetiques pour l'amelioration genetique des cafeiers africains diploi'des.
Travaux et documents, No. 188. ORSTOM, Paris. - Berthaud, J. & Guillaumet, J.L. 1978.
Les cafeiers sauvages en Centrafrique: resultats d'une mission de prospection (janvier-fevrier 1975).
Cafe, Cacao, The 3: 171-186. - Berthaud, J, Guillaumet, J.L, Le Pierres, D. & Lourd, M. 1980.
Les cafeiers sauvages du Kenya: prospection et mise en culture. Cafe, Cacao, The 24: 101-112. - Bridson, D. 1994.
Additionnal notes on Coffea (Rubiaceae) from Tropical East Africa. Kew Bull. 49: 331-342. - Bridson, D. & Verdcourt, B. 1988. Coffea. Pp. 703-723
in: Polhill, R.M. (ed.), Flora of Tropical East Africa, part 2. A.A. Balkema, Rotterdam. - Campa, C., Ballester, J.M., Doulbeau, S., Dussert, S, Hamon, S. & Noirot, M. 2004.
Trigonelline and sucrose diversity in wild Coffea species. Food Chem. 88: 39-43. - Campa, C, Doulbeau, S, Dussert, S, Hamon, S. & Noirot, M. 2005a.
Diversity in bean caffeine content among wild Coffea species: evidence of a discontinuous distribution.
Food Chem. 91: 633-637. - Campa, C, Doulbeau, S, Dussert, S, Hamon, S. & Noirot, M. 2005b.
Qualitative relationship between caffeine and chlorogenic acid contents among wild Coffea species.
Food Chem. 93: 135-139. - Carvalho, A, Medina Filho, H.P, Fazuoli, L.C, Guerreiro Filho, O. & Lima, M.M.A. 1991.
Aspectos geneticos do cafeeiro. Revista Brasil. Genet. 14: 135-183. - Cheek, M, Csiba, L. & Bridson, D. 2002.
A new species of Coffea (Rubiaceae) from western Cameroon. Kew Bull. 57: 675-680. - Chevalier, A. 1947.
Les vrais et les faux cafeiers. Nomenclature et systematique. Encycl. Biol. 28: 115-263. - Combes, M.C, Andrzejewski, S, Anthony, F, Bertrand, B, Rovelli, P., Graziosi, G. & Lashermes, P. 2000.
Characterization of microsatellite loci in Coffea arabica and related coffee species.
Molec. Ecol. 9: 1178-1180. - Coulibaly, I, Noirot, M, Lorieux, M, Charrier, A, Hamon, S. & Louarn, J. 2002.
Introgression of self-compatibility from Coffea heterocalyx to the cultivated species Coffea canephora.
Theor. Appl. Genet. 105: 994-999. - Cros, J. 1996.
Implications phylogenetiques des variations de IADN chloroplastique chez les cafeiers (genres Coffea L. et Psilanthus Hook. f). Doctoral thesis, Theses et Docu ments Microfiches, No. 147. ORSTOM, Paris. - Cros, J, Combes, M.C, Chabrillange, N, Duperray, C, Mon not des Angles, A. & Hamon, S. 1995.
Nuclear content in the subgenus Coffea (Rubiaceae): inter- and intra-specific variation in African species. Canad. J. Bot. 73: 14-20. - Cros, J., Combes, M.C, Trouslot, P, Anthony, F, Hamon, S, Charrier, A. & Lashermes, P. 1998.
Phylogenetic relationships of Coffea species: new evidence based on the chloroplast DNA variation analysis. Molec. Phylog. Evol. 9: 109-117. - Davis, A.P. 2001.
Two new species of Coffea L. (Rubiaceae) from eastern Madagascar. Kew Bull. 56: 479-489. - Davis, A.P, Govaerts, R, Bridson, D.M. & Stoffelen, P. 2006.
An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 152: 465-512. - Davis, A.P. & Mvungi, E.F. 2004.
Two new endangered species of Coffea (Rubiaceae) from the Eastern Arc Mountains (Tanzania) and notes on associated conservation issues. Bot. J. Linn. Soc. 146: 237-245. - Davis, A.P. & Rakotonasolo, F. 2000.
Three new species of Coffea L. (Rubiaceae) from Madagascar. Kew Bull. 55: 405-416. - Davis, A.P. & Rakotonasolo, F. 2001a.
Two new species of Coffea L. (Rubiaceae) from northern Madagascar. Adansonia 23: 337-345. - Davis, A.P. & Rakotonasolo, F. 2001b.
Three new species of Coffea L. (Rubiaceae) from NE Madagascar. Adansonia 23: 137-146. - Davis, A.P. & Rakotonasolo, F. 2003.
New species of Coffea L. (Rubiaceae) from Madagascar. Bot. J. Linn. Soc. 142: 111-118. - Guillaumet, J.L. & Halle, F. 1978.
Echantillonnage du ma teriel recolte en Ethiopie. Bull. Inst. Franc. Cafe Cacao 14: 13-18. - Krug, C.A. & Carvalho, A. 1951.
The genetics of Coffea. Advances Genet. 4: 127-158. - Lashermes, P., Combes, M.C., Robert, J., Trouslot, P., D'Hont, A, Anthony, F. & Charrier, A. 1999.
Molecular characterisation and origin of the Coffea arabica L. genome. Molec. Gen. Genet. 261: 259-266. - Lashermes, P, Combes, M.C, Trouslot, P. & Charrier, A. 1997.
Phylogenetic relationships of coffee tree species {Cof fea L.) as inferred from ITS sequences of nuclear ribosomal DNA. Theor. Appl. Genet. 94: 947-955. - Lashermes, P, Cros, J, Combes, M.C, Trouslot, P, Anthony, F, Hamon, S. & Charrier, A. 1996.
Inheritance and restric tion fragment length polymorphism of chloroplast DNA in the genus Coffea L.
Theor. Appl. Genet. 93: 626-632. - Lebrun, J. 1941. Recherches morphologiques et systematiques sur les cafeiers du Congo. Publ. Inst. Natl. Etude Agron. Congo Beige, Ser. Sei. 11: 1-186.
- Le Pierres, D., Charmetant, P., Yapo, A., Leroy, T., Cou turon, E., Bontems, S. & Tehe, H. 1989.
Les cafeiers sauvages de Cote dTvoire et de Guinee: bilan des missions de prospection effectuees de 1984 á 1987. Pp. 420-428 in: Proceedings of 13th International Scientific Colloquium on Coffee. Association Scientifique Internationale du Cafe, Lausanne. - Louarn J. 1992.
Lafertilite des hybrides interspecifiques et les relations genomiques entre cafeiers diplo'ides d'origine africaine (genre Coffea sous-genre Coffea). Doctoral the sis, Universite Paris-Sud 11, Orsay, France. - Maurin, O., Davis, A.P., Chester, M, Mvungi, E.F, Jaufeer ally-Fakim, Y. & Fay, M.F. 2007.
Towards a phylogeny for Coffea (Rubiaceae): identifying well-supported lineages based on nuclear and plastid DNA sequences. Ann. Bot. 100: 1565-1583. - Namur, C. de, Couturon, E., Sita, P. & Anthony, F. 1987.
Resultats d'une mission de prospection des cafeiers sau vages du Congo. Pp. 397-404 in: Proceedings of 11th International Scientific Colloquium on Coffee. Association Scientifique Internationale du Cafe, Lausanne. - N'Diaye, A., Poncet, V, Louarn, J, Hamon, S. & Noirot, M. 2005.
Genetic differentiation between C. liberica var. liberica and C. liberica var. Dewevrei and comparison with C. canephora. PI. Syst. Evol. 253: 95-104. - Noirot, M, Poncet, V, Barre, P, Hamon, P, Hamon, S. & Kochko, A. de. 2003.
Genome size variations in diploid African Coffea species. Ann. Bot. 92: 709-714. - Poncet, V, Hamon, P, Minier, J, Carasco, C, Hamon, S. & Noirot, M. 2004.
SSR cross-amplification and variation within coffee trees (Coffea spp.). Genome 47: 1071-1081. - Poncet, V, Hamon, P, Sauvage de Saint Marc, M.B., Ber nard, T, Hamon, S. & Noirot, M. 2005.
Base composition of Coffea AFLP sequences and their conservation within the genus. J. Heredity 96: 1-7. - Sonke, B, Ngeumba, C.K. & Davis, A.P. 2006.
A new dwarf Coffea (Rubiaceae) from southern Cameroon. Bot. J. Linn. Soc. 151: 425-430. - Sonke, B. & Stoffelen, P. 2004.
Une nouvelle espece de Cof fea L. (Rubiaceae) du Cameroon, avec quelques notes sur ses affinites avec les especes voisines. Adansonia 26: 153-160. - Stoffelen, P. 1998.
Coffea and Psilanthus (Rubiaceae) in Tropical Africa: A Systematic and Palynological Study, Including a Revision of the West and Central African Species. PhD-thesis, Catholic University Leuven, Leuven, Belgium. - Stoffelen, P, Cheek, M, Bridson, D. & Robbrecht, E. 1997a.
A new species of Coffea and notes on mount Kupe. Kew Bull. 52: 989-994. - Stoffelen, P, Noirot, M., Couturon, E. & Anthony, F. 2008.
A new caffeine-free coffee species of Cameroon. Bot. J. Linn. Soc. 158: 67-72. - Stoffelen, P, Robbrecht, E. & Smets, E. 1996.
Coffea (Rubiaceae) in Cameroon: a new species and a nomen recognized as species. Belg. J. Bot. 129: 71-76. - Stoffelen, P, Robbrecht, E. & Smets, E. 1997b.
Adapted to the rainforest floor: a remarkable new dwarf Coffea (Ru biaceae) from lower Guinea (tropical Africa). Taxon 46: 37-47. - Stoffelen, P, Robbrecht, E. & Smets, E. 1999.
A new species of Coffea (Rubiaceae) from central Africa, with notes on tentative other taxa.
Syst. Geogr. PI. 69: 119-124. - Van der Vossen, H.A.M. 1985.
Coffee selection and breeding. Pp. 48-96
in: Clifford, M.N. & Willson, K.C. (eds.), Coffee Botany, Biochemistry and Production of Beans and Beverage. Croom Helm, London.
'Coffee Botany' 카테고리의 다른 글
| 스테노필라 커피의 플레이버 테이스팅 (9) | 2024.08.30 |
|---|---|
| 스테노필라 커피 (Coffea stenophylla)의 재발견 (3) | 2024.08.29 |
| 리베리카 커피 (2) | 2024.08.15 |
| 카페인-프리 커피나무 C. charrieriana (23) | 2024.08.13 |
| 커피나무와 유전학 서론 (23) | 2024.08.09 |




댓글