
마다가스카르의 코페아 新種 (2003) – 코페아 보헤마렌시스, 코페아 미뉴티플로라, 코페아 모라티
Royal Botanic Gardens, Kew
Coffea vohemarensis → https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:20012584-1
Coffea vohemarensis A.P.Davis & Rakotonas. | Plants of the World Online | Kew Science
The native range of this species is NE. Madagascar. It is a shrub or tree and grows primarily in the wet tropical biome.
powo.science.kew.org
Coffea minutiflora → https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:20012585-1
Coffea minutiflora A.P.Davis & Rakotonas. | Plants of the World Online | Kew Science
The native range of this species is ESE. Madagascar. It is a shrub or tree and grows primarily in the seasonally dry tropical biome.
powo.science.kew.org
Coffea moratii → https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:20012586-1
Coffea moratii J.-F.Leroy ex A.P.Davis & Rakotonas. | Plants of the World Online | Kew Science
The native range of this species is W. Madagascar. It is a shrub or tree and grows primarily in the seasonally dry tropical biome.
powo.science.kew.org
SUMMARY
마다가스카르의 세가지 코페아 新種들이 記述된다:
🌿 코페아 보헤마렌시스 (Coffea vohemarensis),
🌿 코페아 미뉴티플로라 (C. minutiflora),
🌿 코페아 모라티 (C. moratii)
완전한 記述 및 保存 評價가 제시된다.
INTRODUCTION
본고는 마다가스카르의 새로운 코페아 種을 記述하는 일련의 논문 중 마지막 논문이다 (Davis & Rakotonasolo 2000, 2001a, 2001b; Davis, 2001). 이 논문들은 마다가스카르와 코모로의 屬에 대한 지역 개정을 위한 선행 논문을 구성한다. 여기서는 세 가지 새로운 種이 記述되어 마다가스카르의 코페아 種의 총 가지수가 53개가 된다 (코페아 亞屬 Baracoffea 제외). 2000년 이후로 마다가스카르에서 13가지 새로운 코페아 種을 記述했다.
본고에서 사용된 記述的 用語와 規則은 Davis & Rakotonasolo (2000, 2001a, 2001b)에서 제공된 용어와 규칙을 따른다. 마다가스카르의 코페아 형태에 대한 기본 정보와 屬의 간략한 분류학적 역사는 Davis & Rakotonasolo (2001b)에서 제공된다 : 코페아 형태의 자세한 설명은 Davis, Bridson & Rakotonasolo (in press)에 나와 있다.
MATERIAL AND METHODS
Coffea의 식물 표본은 Antananarivo에 있는 Parc de Tsimbazaza의 Département de Botanique (TAN), 파리 국립 자연사 박물관 (P), Antananarivo의 Recherches Forestières et Piscicoles, 그리고 큐 왕립 식물원 (Royal Botanic Gardens, Kew) (K)에서 참조했다. 記述에 나와 있는 측정, 색상 및 기타 세부 정보는 대부분 식물 표본을 기반으로 하지만, 살아 있는 식물과 현장 노트에서 얻은 데이터도 기반으로 한다.
C. moratii의 재배 재료는 Coffee Research Station at Kianjavato (Ministère de la Recherche Scientifique Centre National de Recherche Appliquée au Developpement Rural: FOFIFA)에서 검토되었다; C. vohemarensis는 F. Rakotonasolo가 현장에서 연구했다.
각 종의 보존 상태는 GIS를 사용하여 발생 범위(extent of occurrence)와 점유 면적(area of occupancy)을 계산하고 (J. Moat pers. comm.), IUCN 멸종위기종 목록 카테고리 기준(IUCN, 2001)을 적용하여 평가했다.
DESCRIPTIONS
Coffea vohemarensis
Coffea vohemarensis A.P. Davis & Rakotonas. sp. nov.
Coffea kianjavatensi J.-F. Leroy affinis sed foliis elliptico-oblongo vel late ellipticis-oblongis, rarissime ovatis (nec unquam ellipticis nec elliptico-obovatis nec obovatis nec oblongis) semper chartaceis numquam subcoriaceis, venis lateralibus (venis secundariis) foliorum 5–7-jugis raro 7–9-jugis, stipulis ad apicem acutis (nec apiculo 0.2–0.5 longo ornato) differt.
Typus : Madagascar: Province Antsiranana: Vohemar District: south of Vohemar, Fanambana, Amboay, 190 m, 24.xi.2001 (young fr.), Rakotonasolo RNF 267 (holotypus TAN, isotypi K, TEF).
C. vohemarensis - TREE or SMALL TREE
- TREE or SMALL TREE ⇒ 樹高 c. 7 m, 胸高直徑 d.b.h. c. 7 cm.
- Bark ⇒ not seen.
- BRANCHES ⇒ terete (원통형), 直徑 2.7–3.5 ㎜, 회색~연갈색, 매끄러움.
- BRANCHLETS
⇒ ± terete (원통형), 直徑 1–1.7 ㎜, 회색~연갈색, 매끄러움,
⇒ 미세하고 드문드문 柔毛 (hairs c. 0.05 ㎜ long). - STIPULES
⇒ deltate(삼각주형) to ovate(卵形), 1.5–2.2 × 2–2.5 ㎜,
⇒ subcoriaceous (약간 가죽 같은), 미세하게 柔毛가 있음 (like the branchlets);
⇒ apex → acute (銳頭).
C. vohemarensis - LEAVES
- Petiole (잎꼭지, 葉柄) ⇒ 길이 (0.3–)0.5–0.7 ㎝;
- lamina (잎몸, 葉身)
⇒ elliptic-oblong (타원-길쭉형) to broadly elliptic-oblong, 드물게 ovate (卵形),
⇒ (3.2–)4–7.3(-7.9) × (1.9–)2.5–4.4 ㎝, chartaceous (종이 같은); - Base (잎밑, 葉底) ⇒ attenuate (流底);
- margins (잎 둘레, 葉緣) ⇒ flat (평평);
- Apex (잎끝, 葉先) ⇒ shortly caudate (짧게 尾頭), cauda (꼬리) 길이 0.5–1.2 ㎝;
- abaxial surface (背軸面, 下面):
- midrib (主脈) ⇒ prominent (현저);
- Secondary veins (2차 맥)
⇒ 현저, 7–9 쌍, 30°~45° 각도로 올라감, ± 직선형,
⇒ 둘레에 더 가까이 있는 2차 둘레맥과 결합하여 고리형 둘레맥(hooped intramarginal vein)을 형성; - tertiary venation (3차 맥)
⇒ 현저, ± reticulate (網狀), 2차 맥과 평행인 많은 맥들과 그물 모양 형성; - higher order venation (그 이상 세맥) ⇒ manifest to obscure, ramified (분기됨);
- Adaxial surface (向軸面, 上面) :
- Venation (맥) ⇒ 배축면의 경우보다 덜 명확하게 보임;
- Domatia (巢穴)
⇒ crypt-type (움세포 타입), 눈에 잘 띔, 주맥에 맞닿아 2차 맥의 겨드랑이에 위치,
⇒ 구멍 → slit-like (좁고 기다란 틈), 구경 c. 0.05–0.1 × 0.2–0.7 ㎜,
⇒ 주위 표피는 보통 올라와 있고, 종종 종방향으로 접혀 있음, 無毛.
C. vohemarensis - INFLORESCENCES, CALYCULI, FRUITS
- INFLORESCENCES (꽃차례, 花序)
⇒ 葉腋 당 1개 또는 덜 자주 2개, 花序마다 1개 꽃이 핌, unbranched,
⇒ 길이 3.5–5 ㎜, 과실 발달 중에 신장(elongating),
⇒ 종종 맑은 삼출물로 약하게 덮여 있음. - 화서 축(inflorescence axis) (外萼을 배고 있음) ⇒ 길이 2.2–3 ㎜.
- CALYCULI (外萼)
- 3개, ± cupular (깍정이 모양), ± sessile (착생, 꼭지 없음), 대개 4-열편,
- 약간 가죽 같음, 드문드문 미세하게 柔毛 (hairs 0.05–0.1 ㎜ long),
- 둘레 → 미세하게 섬모 (hairs 0.1–0.2 ㎜ long);
- basal (1st) calyculus
⇒ 때때로 부서지거나 떨어짐, 1–1.7 × 1.7–2.3 ㎜,
⇒ stipular lobes → ± 매우 눌린 卵形 to 거의 切形 (truncate), 0.2–0.5 × 1.4–1.6 ㎜,
⇒ foliar lobes → ± ovate to elliptic, 0.9–1.4 × 0.3–0.4 ㎜; - middle (2nd) calyculus
⇒ 1.1–1.4 × 1.5–2.2 ㎜,
⇒ stipular lobes → depressed ovate, 0.4–0.9 × 1.6–2 ㎜,
⇒ foliar lobes → ± elliptic to ovate, 0.3–0.5 × 0.2–0.3 ㎜; - upper (3rd) calyculus
⇒ 종종 중간 外萼에 의해 가려져 있음, 1–1.2 × 1.1–1.2 ㎜,
⇒ truncate or margin undulate, 많은 분비털(colleters)이 있는 둘레,
⇒ colleters → 좁게 타원형, c. 0.2 ㎜ long, white; - internal surfaces of calyculi
⇒ 분비털들로 성기게 덮여 있음, colleters ± conical, 0.2 ㎜ long. - FLOWERS ⇒ not seen.
- FRUITS ⇒ 매우 불완전하게 알려져 있음, 덜 익은 열매의 꼭지 → 길이 c. 2 ㎜.
C. vohemarensis - Distribution (分布).
- 마다가스카르 북부 Antsiranana 지방의 Vohemar 남쪽과 Sambava 북쪽에 토착 (see Fig. 1).
C. vohemarensis - Habitat and ecology (棲息地 및 生態).
- 다습한 상록림.
- 모래, 석회암, 또는 기반암. 고도 10-20 m.
C. vohemarensis - Phenology (生物季節學).
- 불완전하게 알려져 있음 – 11월에 개화.
C. vohemarensis - Conservation status (保存 狀態).
- Provisional IUCN Red List Category: Endangered (EN B1 a, b).
B1 – extent of occurrence less than 5000 ㎢ (c. 100 ㎢ for C. vohemarensis);
a – severely fragmented and possibly existing at no more than five locations (presently known from only two collections (locations));
b (i–v) – continuing decline inferred. - 확실한 보존 평가가 이뤄질 수 있으려면, Vohemar 地區와 Sambava 地區에서의 추가적인 필드워크가 필요하다.
C. vohemarensis - Other specimens examined.
- E MADAGSCAR: PROVINCE ANTSIRANANA:
- DISTRICT VOHEMAR: south of Vohemar, Fanambana, Amboay, 190 m, 24.xi.2001 (young fr.), Rakotonasolo RNF 269 (K, TAN, TEF);
- DISTRICT SAMBAVA: N of Sambava, 11.xi.1970 (recollected at the Coffee Research Station at Kianjavato by J.-F. Leroy, date of recollection 9.x.1972 (ster.), A. 954 (K, P).
C. vohemarensis - vs. C. kianjavatensis
- Coffea vohemarensis는 C. kianjavatensis와 여러 가지 형태적 유사점을 공유하므로 첫눈에 보기에 매우 비슷하다. 예를 들어, 두 종 모두 거의 같은 크기의 잎을 가지고 있으며, 각각 정점에 뚜렷한 꼬리가 있고, 꽃차례는 작고 매우 단순하다 (無柄, 가지가 없고, 꽃이 1개). Coffea vohemarensis는 C. kianjavatensis와 여러 면에서 다르지만, 주로 잎이 타원형-장방형에서 넓은 타원형 장방형이거나 드물게 난형이고, 항상 흑색이며, 7~9개의 2차 맥이 있고, 3차 맥이 두드러지고 일반적으로 2차 맥과 평행한 방향을 가지고 있다. 턱잎은 정점에서 예리하다. C. kinajavatensis의 잎은 타원형에서 타원형-도란형, 도란형 또는 드물게 ± 장타원형이며, 종이질에서 약간 가죽 같은 질감이며, 보통 5~7쌍의 2차 맥(드물게 8~9개)이 있고, 3차 맥은 약하거나 불분명하고 다소 망상이다. 턱잎은 끝에 apiculum이 있으며, 보통 길이가 약 0.5㎜이다.
- 마다가스카르 북부 및 북서부의 몇가지 코페아 種, 즉 C. dubardii Jum., C. tetragona Jum. & H. Perrier, C. mogenetii Dubard, 그리고 C. heimii J.-F. Leroy는 C. vohemarensis와 유사하게 2차 맥과 평행한 3차 맥을 가지고 있다. 그러나 앞서 언급한 種에서 3차 맥은 훨씬 더 뚜렷하게 평행하고 종종 C. vohemarensis보다 훨씬 더 강하게 나타난다. C. vohemarensis와 대조적으로 C. kianjavetensis의 3차 맥은 약하거나 모호하고 명료하게 網狀이다.
- Coffea vohemarensis와 C. kianjavatensis는 모두 습한 상록수림에서 발생하지만 각 종의 생태는 상당히 다르다.
- Coffea vohemarensis는 해발 200m에서 저지대 숲에서 자라고, 때로는 모래나 석회암 위의 해안 숲에서 자란다.
- Coffea kianjavatensis는 300~600m의 중고도 숲에서 자라지만, 400~600m 사이에서 가장 자주 자라고, 라테라이트 토양에서 발견된다.
- 이 두 종의 분포는 분리되어 있다;
C. vohemarensis는 마다가스카르 북동쪽 Antsiranana 지방에서 자라고,
C. kianjavatensis는 Fianarantsoa 지방의 Kianjavato 근처, Vatovavy 산과 인근 언덕으로 제한되어 있다. - Sambava에서 수집한 표본 A. 951(P!)(1970년 10월 9일, 키안자바토 커피 연구소의 J.-F. Leroy가 FOFIFA로부터 회수)은 C. vohemarensis와 유사하다. 매우 길쭉한 꽃받침을 제외하고는 모든 특징이 새로운 종과 일치한다. 이 형태적 차이가 이 종의 자연적 변이의 일부인지 아니면 다른 요인(예: 재배 유물)인지 확인하기 위해 추가 현장 조사가 필요하다. 우리는 임시로 표본 A. 951을 C. vohemarensis vel aff로 결정했다.
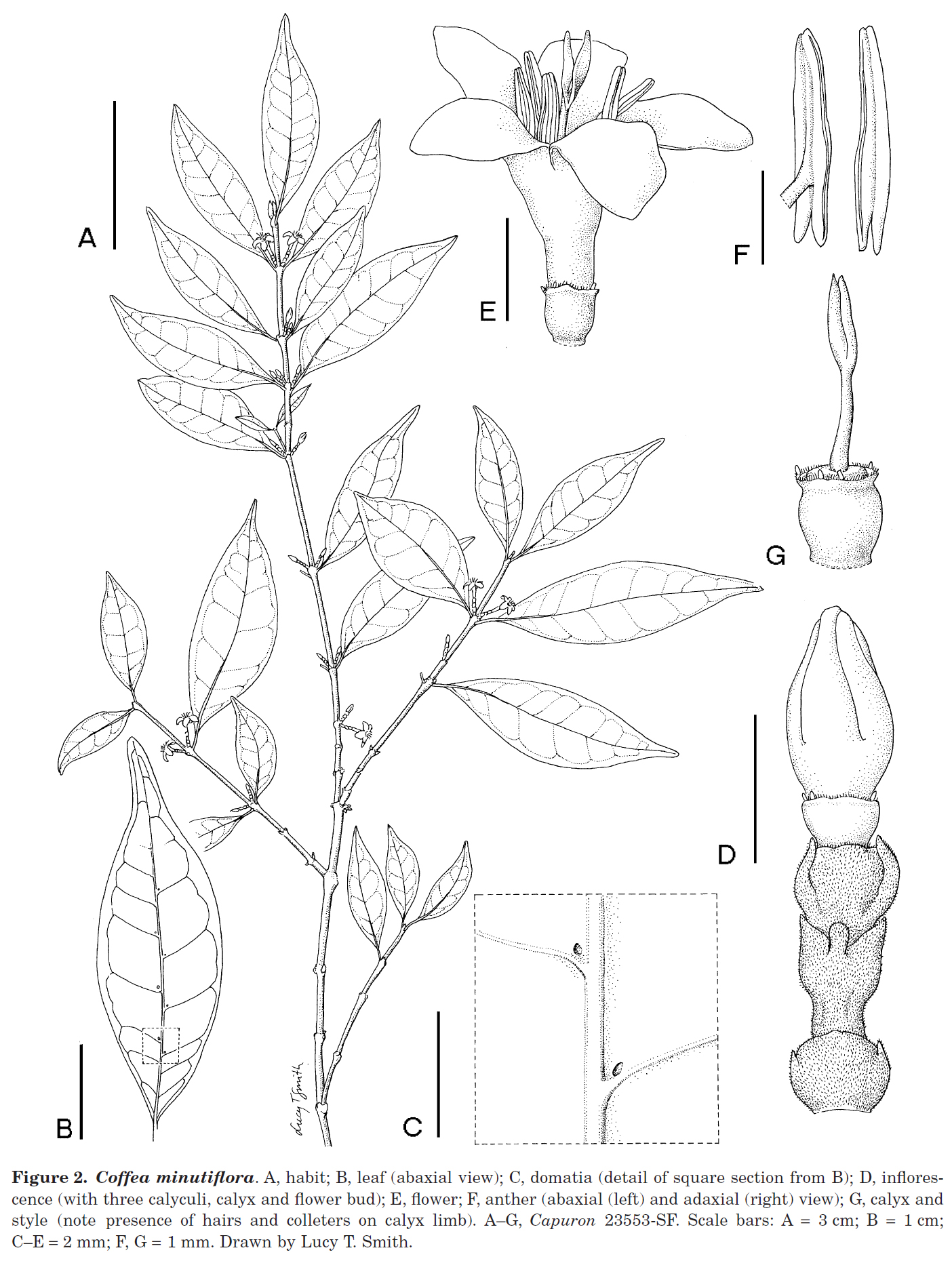
Coffea minutiflora
Coffea minutiflora A.P. Davis & Rakotonas. sp. nov. (Fig. 2)
Coffea vianneyi J.-F. Leroy affinis sed ramulis atque calyculis puberulis (nec glabris), foliis ad apicem breviter caudatis (nec rotundis neque brevissime cau- datis), venis lateralibus (venis secundariis) ad angulum acutissimum 15°-30° tantum (nec 45°-60°) ascendentibus, floribus minoribus, corolla 3.1–5.5 ㎜ longa (nec 6–6.5 ㎜ longo) differt.
Typus : Madagascar: Province Fianarantsoa: Vestige de forêt orientale très dégradée au P.K. 253, de la route Ivohibe-Farafangana, près du village de Beravy, 80–100 m, 16.x.1964 (fl.), Capuron 23553-SF (holotypus P; isotypi BR, K, MO, P, TAN, TEF).
C. mominutiflora - TREE :
- TREE
- 樹高 ⇒ height unknown, 胸高 直徑 ⇒ d.b.h. unknown.
- Bark and branches ⇒ unknown.
- BRANCHLETS
⇒ ± terete, 0.5– 2.5 ㎜ in diam., light grey to medium brown or whitish,
⇒ ± smooth to slightly rough, minutely but densely puberulous (hairs c. 0.05 ㎜ long). - STIPULES
⇒ ± shallowly triangular to very shallowly triangular,
⇒ 0.9–1.2 × 1.4–1.6 ㎜,
⇒ chartaceous to subcoriaceous, puberulous like the branchlets;
⇒ apex broadly acute.
C. mominutiflora - LEAVES :
- petiole (꽃자루, 葉柄) ⇒ (1–)1.5–4 ㎜ long;
- lamina (잎몸, 葉身)
⇒ elliptic-ovate to elliptic, (1.6–)2.5–5.8 × (-0.6)0.8–2.3 ㎝,
⇒ chartaceous (종이 같은) to subcoriaceous (약간 가죽 같은); - base (잎밑, 葉底) ⇒ attenuate (流底) to narrowly cuneate (좁게 楔底) ;
- margins (잎 둘레, 葉緣) ⇒ flat (평평함);
- apex (잎끝, 葉先)⇒ shortly caudate (좁게 尾頭), cauda (꼬리) 길이 0.4–1 ㎝;
- abaxial surface (背軸面, 下面):
- midrib (주맥) ⇒ prominent (현저);
- secondary veins (2차 맥)
⇒ manifest to prominent (뚜렷하고 현저), 6 또는 7 쌍, 15°-30° 각도로 오름,
⇒ ± 직선적, 결합하여 파도모양의 둘레 맥에 고리모양을 형성; - Tertiary venation (3차 맥)
⇒ manifest (뚜렷), ± ramified to reticulate (分枝-網狀); - higher order venation
⇒ manifest to obscure (뚜렷-모호), ± ramified to reticulate (分枝-網狀); - adaxial surface (向軸面, 上面):
- venation ⇒ 배축면의 경우보다 덜 명확하게 뚜렷함 ;
- Domatia (巢穴)
⇒ crypt-type (움-타입), manifest, 2차 맥 겨드랑이 또는 주맥 가장자리에 위치,
⇒ 구멍 → 원형 또는 타원형, 0.1–0.3 × 0.1–0.2 ㎜, 주위 표피=평평, 無毛.
C. mominutiflora - INFLORESCENCES, CALYCULI :
- inflorescence (잎차례, 花序)
⇒ 葉腋(leaf axil) 당 1 또는 2개, 1개 꽃이 핌, unbranched (분기되지 않음),
⇒ 길이 2.5–3.1 ㎜,
⇒ 맑은 삼출물로 연하게 덮여 있거나 거의 덮여 있지 않음; - inflorescence axis (花序 軸) (외악을 배고 있음) ⇒ 길이 1.5–2.5(-3.1) ㎜
- CALYCULI (外萼)
- ⇒ 3개, ± cupular (깍정이 모양), ± sessile (着生, 無柄), 4-lobed,
⇒ 미세하게 조밀하지 않게 柔毛 (hairs c. 0.05 ㎜ long); - basal (1st) calyculus
⇒ usually broken or fallen, 0.5–0.8 × 0.9–1.2 ㎜,
⇒ stipular lobes ± depressed ovate to semi-elliptic, c. 0.3 × 0.6 ㎜,
⇒ foliar lobes broadly ovate, c. 0.6 × 0.5 ㎜; - middle (2nd) calyculus
⇒ 0.5–0.7 ㎜ × 0.9–1.2 ㎜,
⇒ stipular lobes depressed ovate to semi-elliptic, 0.3–0.4 × 0.7 ㎜,
⇒ foliar lobes deltate to triangular, 0.7 × 0.4–0.5 ㎜; - upper (3rd) calyculus
⇒ 1.1–1.4 × 0.9–1.5 ㎜,
⇒ stipular lobes semi- elliptic 0.5 × 0.5–0.7 ㎜;
⇒ foliar lobes ± elliptic to nearly linear, 0.8–0.9 × 0.3–0.4 ㎜; - internal surface of calyculi
⇒ sparsely covered with colleters at base,
⇒ colleters ± triangular, c. 0.1 ㎜ long.
C. mominutiflora - Flowers
- FLOWERS ⇒ 5-갈래 (5-merous);
- Pedicel (꽃자루, 小花柄) ⇒ 0.1–0.2 ㎜ long, or flower almost sessile (거의 무병화).
- Calyx (꽃받침, 花萼) (hypanthium, 꽃턱통)
⇒ ± obconical (0.5–)0.7–1 × 0.8– 1 ㎜; - calyx limb
⇒ undulate to very shallowly 5-lobed,
⇒ margin minutely ciliate (hairs c. 0.05 ㎜ long), and with 3–5 colleters; - colleters ⇒ narrowly cylindrical, c. 0.1 ㎜ long.
- Corolla (꽃부리, 花冠) ⇒ 3.1–5.5 × 5.2–6.5 ㎜;
- corolla tube (꽃부리통) ⇒ 길이1–1.9(-2.6) ㎜;
- corolla lobes (花冠裂片) ⇒ 2–2.6 × 1–1.6 ㎜.
- Stamens (수술, 雄蘂):
- filaments (花絲) ⇒ c. 0.3 ㎜ long;
- Anthers (꽃밥, 花葯)⇒ 2.5–3.2 ㎜ long.
- Ovary (씨방): disc low domed, entire.
- Style (암술대) ⇒ (2.5–)3.5–5.3 ㎜ long;
- stigma lobes (암술머리 편)⇒ 0.8–1.1 ㎜ long.
- FRUITS ⇒ not known.
C. mominutiflora - Distribution (分布).
- 마다가스카르 남동부 피아나란초아 (Fianarantsoa) 지방에 토착.
C. mominutiflora - Habitat and ecology (棲息地 및 生態).
- Humid, evergreen forest. On basement rocks. Altitude 80–100 m.
다습한 상록림. 기반암. 고도 80-100 m.
C. mominutiflora - Phenology (生物季節學).
- 불완전하게 알려져 있음 – 10월에 개화
C. mominutiflora - Conservation status (保存 狀態).
- IUCN Red List Category: Data Deficient (DD).
- 이 종의 보호상태 평가를 위해서는, Ivohibe와 Farafangana 사이의 지역 그리고 덜 수집된 마다가스카르 남동부 다습림에서의 서베이 작업이 필요하다.
- Coffea minutiflora는 현재 한 컬렉션에서만 알려져 있다.
C. mominutiflora - Specimens examined (檢討 標本).
- Only known from the type specimen.
C. mominutiflora - vs. Coffea vianneyi
- 파리(P)와 안타나나리보(TEF)의 식물 표본실에서 Capuron 23553-SF 표본은 C. vianneyi로 식별되지만, 이 자료를 자세히 살펴보면 이것이 별개의 종을 나타내는 것으로 보이며, 여기서는 C. minutiflora로 기술했다.
- Coffea minutiflora는
柔毛가 있는 작은 가지와 外萼(calyculi), 그리고 더 작은 꽃부리(3.1–5.5 × 5.2–6.5 ㎜; 꽃부리통 길이 1–1.9(-2.6) ㎜; 꽃부리 열편 2–2.6 × 1–1.6 ㎜)를 가지고 있고, 2차 맥은 주맥에 15°-30° 각도로 올라가고, 잎 끝은 짧지만 뚜렷하게 꼬리 모양, 게다가 calyx limb에는 미세한 섬모가 있고 3–5개의 분비털들로 둘러싸여 있다는 점에서 C. vianneyi와 다르다. - Coffea vianneyi는
무모의 잔가지들과 外萼 (윗부분의 꽃받침은 미세한 유두 모양(papillate))을 가지고 있으며, 꽃부리가 더 크고 (6–6.5 × 7–8 ㎜; 꽃부리 통 길이 0.8–1.2 ㎜; 꽃부리 열편 4.5–5 × 1.6–1.9 ㎜), 2차 맥은 주맥에서 45°-60° 각도로 올라가고, 잎 끝은 둥글거나 때로는 매우 짧게 꼬리 모양(尾頭)이며, calyx limb에는 털과 분비털들이 없다. - 이 종의 꽃은 아마도 모든 코페아 종 중에서 가장 작으며, 따라서 우리는 이 종을 C. minutiflora라고 명명하기로 했다. 현재 C. minutiflora는 Ivohibe와 Farafangana (province Fianarantsoa) 사이의 작은 숲 부분에서 수집한 단일 수집(Capuron 23553-SF)에서 알려져 있다. 이 종은 현재 단일 gathering (여러 중복이 있는 하나의 컬렉션)에서만 알려져 있지만, 우리는 이 종이 의심할 여지 없이 별개의 구별적인 것이라고 믿는다.
Coffea moratii
Coffea moratii J.-F. Leroy ex A.P. Davis & Rakotonas. sp. nov. (Fig. 3)
Coffea moratii J.-F. Leroy, in sched (P).
Coffea sakarahae J.-F. Leroy affinis sed foliis ellipticis vel late ellipticis vel ovatis vel late ovatis vel rarissime fere orbicularibus (nec obovatis nec oblanceolatis nec ellipticis nec anguste tantum ellipticis) (4.4–)5.5–7.7 × (2.3–)2.9–4.5(-5.3) cm (nec (1.2–)2.1–5(-5.2) × (0.5–) 0.7–1.8(-2.2) cm), venis lateralibus (venis secundaris) foliorum semper 6–9-jugis (nec 4–5(6–7)-jugis), floribus 7-meris (nec 5-meris), corollis 15–18 ¥ 13–18 ㎜ (nec 4.5–6 × 4–6 ㎜), corollae tubo 7.5–9 ㎜ longo (nec 2.5–3.5 ㎜) differt.
Typus : Madagascar: Province Tulear: RN 9 [Réserve Tsingy de Bemaraha], recollected from the Coffee Research Centre at Kianjavato (Kianjavato acc. no. A. 740), date of recollection 27.xi.1999 (fl.), Davis & Rakotonasolo 2326 (holotypus K; isotypi P, MO, TAN, TEF).
C. moratii - TREE or TREELET
- TREE or TREELET, ⇒ 樹高 c. 1.8–4.5 m high, 胸高直徑 d.b.h. c. 3–6 ㎝.
- Bark ⇒ ± 매끄러움~약간 거침 smooth to slightly rough, 연갈색~진갈색.
- BRANCHES ⇒ terete (가는 원통형), 직경 6–10 ㎜, 갈색~연갈색, 매끄러움~균열적(fissured).
- BRANCHLETS
⇒ ± terete, 직경 (1.5–)2–4.2(-5.5) ㎜, 연갈색~갈색,
⇒ 매끄러움~거침, 가끔 벗겨진 껍질이 있음. - STIPULES
⇒ 눌린 난형~얕게 삼각형, (1.5–)0.9–1.2 × 2–3 ㎜, 종이 같음(chartaceous), 無毛;
⇒ 턱잎끝 → 넓은 銳頭(acute)~鈍頭(obtuse).
C. moratii - LEAVES
- LEAVES:
- Petiole (잎꼭지, 葉柄) ⇒ 길이 (2.5–)3–5 ㎜;
- lamina (잎몸, 葉身)
⇒ 타원형~넓게 타원형, 또는 卵形~넓게 卵形, 드물게 거의 圓形(orbicular),
⇒ (4.4–)5.5–7.7 × (2.3–)2.9–4.5(-5.3) cm,
⇒ 약간 가죽 같음(subcoriaceous); - base (잎밑, 葉底)
⇒ 流底(attenuate), 또는 楔底(cuneate)~圓底(rounded),
⇒ 종종 약간 줄기 아래로 뻗음(decurrent); - margins (잎둘레, 葉緣) ⇒ 약간 反曲(subrevolute);
- apex (잎끝, 葉先) ⇒ 銳頭(acute)~넓게 銳頭(acute);
- abaxial surface (背軸面, 下面):
- Midrib (主脈) ⇒ 顯著 (prominent);
- secondary veins (2次脈)
⇒ 뚜렷~顯著, 6–9쌍, 약 45° 각도로 올라감,
⇒ 약간 곡선 및 종종 구부러짐(crooked) (보통 맥이 소혈과 만나는 곳에서) ~ ± 직선적,
⇒ 둘레에 더 가까운 분명한 둘레맥과 결합하여 고리형 둘레맥을 형성; - tertiary venation (3次脈)
⇒ 뚜렷~모호, 網狀 (reticulate); - higher order venation (그 이상 세맥) ⇒ 뚜렷~모호, ± 網狀 (reticulate)~分岐(ramified);
- adaxial surface (向軸面, 上面):
- Venation (맥) ⇒ 배축면의 경우보다 덜 명확하게 또는 더 명확하게 드러남;
- domatia (巢穴)
⇒ 움 타입(crypt type), 현저,
⇒ 2次脈 겨드랑이에 위치하지 않고 주맥으로부터 2차맥을 따라 (2–)3– 6 ㎜ 정도에 위치한다,
⇒ 구멍 ± 타원형, 구경 0.2–0.5 × 0.2–0.3 ㎜,
⇒ 둘레 및 주위 표피는 약간 올라와 있음,
⇒ 무모 또는 적은 수의 갈색 털들이 둘레 내에 있음 (c. 0.2 ㎜ long),
⇒ 향축면에서의 소혈은 더 현저하고, 작은 둥근~타원형 농포처럼 눈에 띔.
C. moratii - INFLORESCENCES, CALYCULI
- INFLORESCENCES (꽃차례, 花序)
⇒ 첫번째 또는 두번째 잎 쌍의 葉腋에 있음,
⇒ 葉腋 당 1-3개, 1개의 꽃이 핌, 가지는 뻗지 않음,
⇒ 길이 2–7.6 ㎜, 과실 발달 중에는 약간 늘어남,
⇒ 맑은 渗出物으로 덮여 있지 않거나 매우 약간 덮여 있음.
⇒ 화서축 (외악을 배고 있음) → 길이 2–5 ㎜. - CALYCULI (外萼)
⇒ 2 또는 3개 (첫번째 외악은 없거나 금방 떨어짐),
⇒ ± 깍정이 모양(cupular), ± sessile (着生),
⇒ 4-lobed (그러나 foliar lobes는 종종 떨어지며, 그러면 외악은 2-편으로 보임),
⇒ chartaceous(종이질), glabrous(無毛),
⇒ 둘레 → 無毛, hyaline (유리질, 투명체); - basal (1st or 2nd)
⇒ 2–4 × 2–2.5 ㎜,
⇒ stipular lobes hemi-elliptic to ± deltate, 1.2–2.1 × 2 ㎜,
⇒ foliar lobes triangular to shallowly triangular, 0.3–0.5 × 0.3–0.5 ㎜; - upper (2nd or 3rd) calyculus
⇒ 3–4.5(-5.5) × 2–2.5 ㎜,
⇒ stipular lobes hemi-elliptic to ± deltate, 1.5–2 × 2–2.2 ㎜,
⇒ foliar lobes absent (?fallen) or narrowly elliptic to oblanceolate,
1.8–1.9 × 0.6–0.7 ㎜, or the same as the foliar lobes of the lower calyculus; - internal surfaces of calyculi
⇒ 분비털들로 덮여 있음 (특히 basal의 ⅔에서),
⇒ 분비털(colleters)→ ± narrowly conical, 길이 0.2–0.3 ㎜.
C. moratii - FLOWERS
- FLOWERS 꽃 ⇒ (5–)6–7-merous;
- Pedicel (꽃자루, 花柄) ⇒ 없거나 매우 짧음, 길이 0–0.2 ㎜.
- Calyx (꽃받침, 花萼) (hypanthium, 꽃턱통) ⇒ ± obovoid (타원체), 1.9–2.5 × 2.3–2.5 ㎜;
- calyx limb (꽃받침 테)
⇒ truncate (截形), 불룩한 고리모양의 테(annulate rim), 디스크보다 분명히 더 짧음, 둘레는 무모. - Corolla (꽃부리, 花冠) ⇒ 15–18 × 13–18 ㎜;
- corolla tube (꽃부리 통) ⇒ 7.5–9 ㎜ long;
- corolla lobes (화관열편) ⇒ 6–7.5 × 3.1–3.5 ㎜.
- Stamens (수술, 雄蘂):
- Filaments (花絲) ⇒ 1–1.2 ㎜ long;
- Anthers (꽃밥, 花葯) ⇒ 4.8–5.5 ㎜ long.
- Ovary (씨방) : disc ⇒ 현저, low domed, entire.
- Style (암술) ⇒ 10.1–13.6 ㎜ long;
stigma lobes (암술머리 편)⇒ 3.8–4.6 ㎜ long. - FRUITS (과실)
⇒ 불완전하게 알려져 있음.
⇒ 아주 어린 열매만 보임, ± 타원체~타원-도란형(obovoid), 5–9 × 4 ㎜. - Seeds ⇒ not seen.
C. moratii - Distribution (分布).
- 마다가스카르 서부에 토착 (Endemic to western Madagascar);
- 아마도 Réserve Tsingy de Bemaraha 및 인근의 적합서식지에 제한적일 것이다.
C. moratii - Habitat and ecology (棲息地 및 生態).
- Seasonally dry deciduous forest (containing some evergreen species).
- Tsingy limestone and possibly other limestone formations.
- Altitude 50–100 m, and possibly up to 600 m.
C. moratii - Phenology (生物季節學).
- 불완전하게 알려져 있음 – 10월과 11월에 개화.
C. moratii - Conservation status (保存 狀態).
- Provisional IUCN Red List Category: Endangered (EN B1a, b, and B2 a, b).
- B1 – total extent of occurrence less than 5000 ㎢ (c. 1360 ㎢ for C. moratii);
- B2 – area of occupancy less than 500 ㎢ (c. 40 ㎢);
- a – severely fragmented, and possibly existing at no more than five locations (presently known from only three collections (locations));
- b (i–v) – continuing decline inferred.
- C. moratii의 잠정적인 점유 지역과 발생 범위가 추가적인 식물 조사 작업으로 증가할 가능성이 있으므로 임시 보존 평가가 주어진다. 예를 들어, Réserve Tsingy de Bemaraha에서 이 종의 이용 가능한 서식지는 상당하다.
C. moratii - Specimens examined (檢討 標本).
- W MADAGSCAR: PROVINCE TULEAR:
- Antsalova, Bekopaka, Res. no. 9 [Réserve Tsingy de Bemaraha], x.1964 (fl.), Morat 876 (K, P); Reserve no. 9 d’Antsalova [Réserve Tsingy de Bemaraha], sur la route de Tsiandro, à 3 km à l’entrée de la réserve, 6.x.1968 (ster.), Guillaumet 2268 (K, P);
- dans les couloirs des Tsingy à 70 km de Maintirano sur le route de Morafenobe, xi.1968 (ster.), Guillaumet 2261a (K, P);
- RN 9 [Réserve Tsingy de Bemaraha], recol- lected from the Coffee Research Centre at Kianjavato (Kianjavato acc. no. A. 740), date of recollection 27.xi.1999 (fl.), Davis & Rakotonasolo 2326 (holotype K; isotypes P, MO, TAN, TEF).
C. moratii - vs. C. sakarahae
- Coffea moratii는 C. sakarahae와 많은 특징을 공유하며, 가까운 공통 조상을 공유했을 가능성이 높다. 두 종 모두 잎의 주맥에서 어느 정도 떨어진 곳에 도마티아가 있는 잎을 가지고 있다. 대부분의 Coffea 종에서 도마티아는 주맥에 맞닿아 있거나 매우 가까이에 있다. 이 종들의 꽃차례는 싹의 끝 부분, 1번째, 2번째 또는 3번째 잎 쌍의 겨드랑이에서만 발견되며(Fig. 2 참조), 外萼은 매우 작은 엽편(foliar lobes)을 가지고 있다.
- Coffea moratii는 C. sakarahae와 달리 잎이 뚜렷이 더 크고, 보통 5.5~7.7 × 2.9~4.5cm, 楕圓形~넓은 楕圓形 또는 卵形~넓은 卵形이며, 6~9쌍의 2차 맥이 있다. 또한 꽃은 6瓣 또는 7瓣 (드물게 5瓣)이고, 꽃부리는 15–18×13–18㎜이다. C. sakarahae의 잎은 보통 2.1–5×0.7–1.8cm이고, 倒卵形에서 披針形 또는 楕圓形에서 좁은 楕圓形이며, 4–5쌍 또는 때로는 6–7쌍의 2차 맥이 있다. 꽃은 5瓣이고, 꽃부리는 4.5–6×4–6㎜이다.
- C. moratii와 C. sakarahae는 모두 계절적으로 건조한 숲, 반 상록림(낙엽수 종 포함) 또는 주로 낙엽수(deciduous)림에서 발견된다. 그러나 C. moratii는 Tsingy 석회암에 국한되는 반면, C. sakarahae는 주로 모래와 기반 암석(basement rocks)에서 발견된다.
- 이 두 종의 분포는 겹치지 않는다. C. sakarahae는 주로 Site d’Internet Biologique de Zombitse (Sakaraha 숲 포함)의 삼림 지역과 Ihosy (Fianarantsoa 및 Tulear 지방) 반경 20km 이내의 삼림 지역에서 발견된다. C. moratii는 Bekopaka 근처의 Tsingy de Bemaraha 보호구역에서 발견되며, 아마도 보호구역에 인접한 비슷한 서식지 지역에서 발견된다.
- Coffea Moratii는 프랑스 파리 국립자연사박물관의 Laboratoire de Phanérogamie 관장인 P. Morat 교수의 이름을 따서 명명되었다.

REFERENCES
- Davis AP. 2001.
Two new species of Coffea (Rubiaceae) from eastern Madagascar.
Kew Bulletin 56: 479–489. - Davis AP, Bridson DM, Rakotonasolo F. In press.
A reexamination of Coffea subgenus Baracoffea and comments on the morphology and classification of Coffea and Psilanthus (Rubiaceae-Coffeeae).
Annals of the Missouri Botanical Garden. in press. - Davis AP, Rakotonasolo F. 2000.
Three new species of Cof- fea (Rubiaceae) from Madagascar. Kew Bulletin 55: 405–416. - Davis AP, Rakotonasolo F. 2001a.
Three new species of Cof- fea from northern Madagascar.
Adansonia Sér. 3 (23): 137–146. - Davis AP, Rakotonasolo F. 2001b.
Two new species of Coffea (Rubiaceae) from northern Madagascar: C. ankaranensis and C. sambavensis.
Adansonia Sér. 3 (23): 337–345. - Humbert H. 1955.
Les territoires phytogéographiques de Madagascar. Leur cartographie.
Colloque sur les Régions Ecologiques du Globe, Paris, 1954. Année Biologique 31: 195–204. - IUCN. 2001. IUCN Red List Categories: Version 3.1. IUCN Species Survival Commission, Gland, Switzerland and Cambridge, UK.
'Coffee Botany' 카테고리의 다른 글
| 코페아 앤카라넨시스, 코페아 샘바벤시스 (2) | 2024.10.14 |
|---|---|
| 코페아 리토랄리스, 코페아 맥퍼슨니, 코페아 라치마망게 (0) | 2024.10.13 |
| 코페아 라코토나소로이, 코페아 몬티스-새크리 (0) | 2024.09.29 |
| 코페아 마놈벤시스, 코페아 리아우디, 코페아 리로이 - 신종코페아 3가지 (2000) (2) | 2024.09.28 |
| 코페아 퍼스튤라타, 코페아 마이크로두바르디, 코페아 루피콜라, 코페아 칼만데리, 코페아 다라이넨시스, 코페아 칼로비노넨시스 (0) | 2024.09.16 |




댓글