
ABSTRACT
마다가스카르 北部 (Antsiranana Province)로부터의 새로운 코페아 2種들이 記述된다:
🌿 코페아 앤카라넨시스 (C. ankaranensis)
🌿 코페아 샘바벤시스 (C. sambavensis)
이 2種들이 각각 C. perrieri와 C. millotii와 비교된다.
각 種의 保存 狀態가 IUCN Red List Categories를 사용하여 요약된다.
Coffea ankaranensis https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:20003333-1
Coffea ankaranensis J.-F.Leroy ex A.P.Davis & Rakotonas. | Plants of the World Online | Kew Science
The native range of this species is N. Madagascar. It is a tree and grows primarily in the wet tropical biome.
powo.science.kew.org
Coffea sambavensis https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:20003334-1
INTRODUCTION
마다가스카르에서 발견되는 45種 정도의 코페아 種들 가운데 26種이 LEROY에 의해 記述되었다 (예: 1961, 1962a, 1962b, 1972a, 1972b). 그는 또한 파리 국립 자연사 박물관(P)의 식물 표본관에 보관된 표본에 쓰여진 대로 여러 種에 임시 이름을 붙였다. 이러한 이름 중 다수는 현재 유효하게 발표되었지만 (예: DAVIS & RAKOTONASOLO 2000, 2001, DAVIS 2001), 원래 별명 중 일부는 사용되지 않았다. 본고에서 우리는 LEROY의 임시 種들 가운데 2가지 種, 즉 C. ankaranensis와 C. sambavensis에 공식적으로 이름을 붙였다. 이 2가지 種은 밀접한 관련이 없지만, 둘 다 중간 크기의 잎을 가지고 있으며 안치라나나(Antsiranana) 지방에서 유래되었기 때문에 여기에서는 함께 記述한다.
Coffea는 세 개의 亞屬들(subgenera)로 구성되어 있다:
- Subg. Coffea (약 90種),
- Subg. Psilanthopsis (A. Chev.) J.-F. Leroy (1種),
- Subg. Baracoffea (J.-F. Leroy) J.-F. Leroy (약 4種).
그러나 BRIDSON (1994)에 따르면 Psilanthopsis 亞屬은 인정받을 만 하지 않다; 이 분류군의 단일 種인 C. kapakata (A. Chev.) Bridson은 Coffea 亞屬으로 옮겨야 한다. 그리고 DAVIS, BRIDSON & RAKOTONASOLO (in prep.)의 형태학적 연구에 따르면 Baracoffea 亞屬은 Coffea 亞屬 보다 Psilanthus 屬에 더 가깝다.
코페아 앤카라네시스(Coffea ankaranesis)와 코페아 샘바벤시스(C. sambavensis)는 코페아 亞屬의 멤버이다.
▣ Coffea subg.
Coffea는 屬의 기준 種인 C. arabica에 의해 예시된다.
Coffea 亞屬의 구성원은 다음과 같은 두드러진 특징의 조합으로 특징지어질 수 있다.
- 잎은 거의 항상 常綠樹이다.
- 꽃차례는 葉腋에 있고, 外萼(calyculi)은 포-같은 엽편들(bract-like lobes)을 가지고 있다.
- 싹의 구조는 개화에 의해 바뀌지 않는데, 꽃차례는 결정되어 있기 때문이다 (relay axes를 생성하지 않음).
- 꽃받침(calyx)은 디스크보다 짧거나 약간 더 길다.
- 꽃부리(corolla)은 흰색이거나 드물게 분홍색이고, 꽃부리 관(corolla tube)은 보통 花冠裂片(corolla lobes)보다 짧거나 약간 더 길다;
- 수술(stamens)과 암술대(style)는 튀어나온다.
- 열매는 核果(drupes)이고, 씨앗 두 개 또는 때로는 씨앗 하나가 들어 있으며,
각 씨앗은 向軸(안쪽)面에 깊은 陷入(invagination)이 있다 (즉, "커피콩").
▣ Baracoffea 亞屬
Coffea 屬의 Baracoffea 亞屬은 Coffea 亞屬과 많은 특징을 공유하지만, 이 亞屬 사이에는 몇 가지 차이점이 있다.
- 잎은 낙엽성(deciduous)이다.
- 꽃차례는 겨드랑이에 있거나 또는 끝에 있으며,
(가장 위쪽) 外萼(calyculi)은 잎처럼 생긴 엽(leaf-like lobes)을 가지고 있다. - 꽃이 피면 싹(shoot)의 구조가 바뀌는데, 꽃차례가 불확정적이기 때문이다(relay axes를 생성).
- 꽃부리 통(corolla tube)은 항상 花冠裂片(corolla lobes) 보다 훨씬 길고, 그리고 수술은 부분적으로만 드러난다.
▣ Calyculi (外萼)은
- Coffea 種과 Coffeeae 屬을 식별하는데 중요한 구조이며, 여기서 그 구조에 대한 정보를 제공할 필요가 있다.
- Calyculi (複數)과 calyculus (單數)라는 용어는 ROBBRECHT(1978), STOFFELEN(1998) 및 DAVIS 2000을 포함한 여러 Rubiaceae 연구자들이 사용했다.
- Calyculi (外萼)은
꽃, 그리고 꽃자루(pedicel(s))와 꽃차례 가지와 같은 꽃차례의 다른 부분들을 덮는
苞葉 같은 구조(bract-like structures)이다. - 일반적으로 꽃차례 하나에 外萼이 3개 있지만,
종마다 그리고 때로는 종 내에서도 그 숫자가 다르며,
최소 2개에서 최대 25개, 또는 그 이상으로 상당히 다를 수 있다. - 각 外萼에는 4개의 엽(lobes)이 있는데,
2개는 턱잎에서 유래한 턱잎 엽(stipular lobes)이라고 하고,
2개는 잎에서 유래한 잎 엽(foliar lobes)이라고 한다. - 때때로 그 엽편들은 비교적 크고 눈에 띄며, 그것의 유래가 stipular인지 또는 foliar인지가 분명하거나, 또는 작고 다소 중요하지 않을 수도 있다.
- 外萼은 엽편(lobes) 아래에 융합되어 다소 컵 모양 또는 관상 구조를 형성한다.
Coffea 屬을 다루는 최근의 논문들은 없으며, 마지막이자 가장 중요한 것이 CHEVALIER의 논문들 (예: 1942, 1947)이지만, 이 저작들은 이제 매우 오래되었다.
그러나 여러 지역적 개정들이 있는데, 특히 동아프리카에 대한 BRIDSON(1988)과 중부 및 서아프리카에 대한 STOFFELEN (1998)의 개정이 있다.
마다가스카르, 코모로 제도, 마스카렌 제도의 Coffea에 대한 지역적 취급이 우리(DAVIS & RAKOTONASOLO, 준비 중)에 의해 준비 중이다.
MATERIAL AND METHODS
- Coffea의 식물 표본 자료는
Département de Botanique, Parc de Tsimbazaza, Antananarivo (TAN),
Muséum National d’Histoire Naturelle, Paris (P),
Recherches Forestières et Piscicoles, Antananarivo (TEF),
Royal Botanic Gardens, Kew (K)에서 참조했다. - 記述에 나와 있는 측정, 색상 및 기타 세부 정보는 대부분 식물 표본을 기반으로 하지만, 살아 있는 식물과 현장 노트에서 얻은 데이터도 기반으로 한다.
- C. Ankaranensis와 C. sambavensis의 재배 자료는
Kianjavato (FOFIFA)의 Coffee Research Station에서 검토했다. - C. Sambavensis는 F. RAKOTONASOLO가 야생에서 연구했다.
- 각 種의 保存 狀態는
GIS (J. MOAT 개인 커뮤니케이션)를 사용하여 extent of occurrence (발생 범위)를 계산하고 IUCN 멸종위기종 목록 범주 기준(IUCN 2001)을 적용하여 평가했다.
THE NEW SPECIES
Coffea ankaranensis
Coffea ankaranensis J.-F. Leroy ex A.P. Davis & Rakotonas., sp. nov.
Coffeae perrieri Drake ex Jum. & H. Perrier affinis sed foliis latioribus plerumque ovalibus vel obovatis vel late ellipticis (nec ellipticis nec elliptici-oblongis nec anguste ellipticis nec elliptici-oblanceolatis), calyculis pubescentibus vel villosis (nec puberulis), floribus majoribus [13.5-17 × 12-14 mm nec 10-13(-16) × 6-14(-20) mm] et fructibus majoribus (2.3-2.5 × 1.1-1.3 cm nec 1.1-1.2 × 0.9-1 cm) distinguenda.
TYPUS. — Capuron 23166-SF, Madagascar, Ouest (Nord): plateau de l’Ankarana, massif d’Antsandoko, au Sud d’Antsandoko, au Sud d’Ambondromifehy, 22 Dec. 1963, fl. (holo-, P!; iso-, K!, TEF!).
C. ankaranensis - TREE or SMALL TREE
- Tree or small tree, ⇒ 樹高 (3-)4-10 m, 胸高 直徑 dbh (3.5-)5-9(-10) ㎝.
- Bark ⇒ 암갈색~회갈색, 매끄러움(smooth).
- Branches ⇒ 원통형(terete), 직경 6-10 ㎜, 연회색 또는 연갈색, 또는 희끄무레한 색, 매끄러움.
- Branchlets
⇒ ± 원통형 (terete), 직경 2-5 ㎜, 연회색~희끄무레한 색 또는 연갈색,
⇒ 매끄러움~약간 거침, 가끔 말랐을 때 갈라짐(splitting), 無毛(glabrous). - Stipules
⇒ 일찍 떨어짐(caducous), 델타형(deltate)~삼각형(triangular), 1.6-2 × 1.8-3 ㎜,
⇒ 약간 가죽 같음(subcoriaceous)~± 나무 같음(woody),
⇒ 보통 작은 구멍이 있는 질감(pitted texture), 無毛(glabrous),
⇒ 둘레 → 無毛(glabrous); - apex (턱잎 끝) ⇒ 銳頭(acute)~짧게 뾰족, apiculum c. 0.5 ㎜ long.
C. ankaranensis - Leaves
- Petioles (잎자루, 葉柄) ⇒ 0.4-0.7 ㎝ long;
- Leaf-blades (잎새, 葉片)
⇒ 광타원형(oval), 도란형(obovate), 또는 넓게 타원형(broadly elliptic)~매우 넓게 타원형, 드물게 타원형,
⇒ (4.7-)5.4-12 × (2.7-)4.4-6.2 ㎝, 약간 가죽 같음(subcoriaceous); - Base (잎밑, 葉底) ⇒ 流底(attenuate);
- Margins (잎둘레, 葉緣) ⇒ 약간 反曲(subrevolute)~反曲(revolute);
- Apex (잎끝, 葉先) ⇒ 圓頭(rounded)~鈍頭(obtuse) 또는 약간 漸尖頭(subacuminate);
- Midrib (主脈) ⇒ prominent;
- Abaxial surface (背軸面, 下面):
- Midrib (主脈) ⇒ 현저(prominent);
- Secondary veins (2次脈)
⇒ 뚜렷(manifest), (6-)8-10 쌍, 약 45° 각도로 올라감, 직선적~약간 곡선적,
⇒ 결합하여 고리모양의 둘레맥 (hooped intramarginal vein)을 형성; - Tertiary venation (3次脈) ⇒ 약함~모호, 網狀(reticulate);
- Higher order venation (그 이상 細脈) ⇒ 보이지 않음;
- Adaxial surface (向軸面, 上面):
- Venation (脈) ⇒ 배축면보다 덜 명확하게 뚜렷함;
- Domatia (巢穴)
⇒ 움 타입(crypt type), 희미(indistinct)~현저(prominent),
⇒ 2차 맥의 겨드랑이에 위치, 주맥의 반대쪽 또는 가장자리,
⇒ 원형 구멍, 口徑 0.5-0.7 ㎜, 또는 卵形(oval)~좁고 기다란 틈 같음(slit-like), c. 0.1 × 0.2 ㎜,
⇒ 둘레 → 종종 종이 같음(chartaceous) 또는 darkened, 그리고 약간 부풀어 오름, 無毛;
⇒ 향축면에서 不可示~희미.
C. ankaranensis - Inflorescences, Calyculi
- Inflorescences (꽃차례, 花序)
- 葉腋당 1개, 4-6개의 꽃이 핌, ± umbellate (繖形),
- unbranched 또는 매우 짧은 가지 1개도 있음 (c. 0.2 ㎜ long),
- 길이 10-15 ㎜, 과실 발육 중에 신장,
渗出物로 덮여 있지 않거나 거의 덮여 있지 않음; - inflorescence axis (花序軸) (bearing calyculi) ⇒ 길이 5-11 ㎜.
- Calyculi (外萼)
- 4개, 기저 외악은 종종 떨어짐, ± sessile (착생, 무병), 약간 가죽 같음(subcoriaceous),
연모 있음(pubescent)~ 絨毛 있음(villous), - 둘레 → 털이 많음 (0.3-0.6 ㎜ long), 분비털들(colleters)이 가끔 있음;
- basal (1st) calyculus
⇒ 2.5-3 × 3.1-3.5 ㎜,
⇒ stipular lobes → broadly ovate, 2.5-3 × 2.3-2.5 ㎜,
⇒ foliar lobes → ovate-elliptic, 2-2.3 × 1.1-1.3 ㎜; - middle (2nd) calyculus
⇒ 2.3-3.5 × 4-4.3 ㎜,
⇒ stipular lobes → broadly elliptic to obovate, 3-3.2 × 2.4-2.6 ㎜,
⇒ foliar lobes → ± ligulate(舌狀) to elliptic, 1-1.3 × 0.4-0.5 ㎜; - middle calyculus (3rd)
⇒ 2.5-3.3 × 4-4.5 ㎜,
⇒ stipular lobes → ± square to transversely oblong, 2.7-2.8 × 2.6-2.8 ㎜,
⇒ foliar lobes → ovate-elliptic, 2.2-2.3 × 1.1-1.3 ㎜; - upper calyculus (4th)
⇒ cupular to ± tubular, 1.1-1.7 × 2.5-2.8 ㎜, not lobed; - 외악 내면
⇒ 분비털로 조밀하게 덮여 있고, 특히 기저 부분에 덮여 있음,
⇒ 가늘고 희끄무레한~반투명한 털들이 섞여 있음 (0.2-0.4 ㎜ long); - 분비털들 → 좁은 타원체~원추형, 길이 0.2-0.3 ㎜, 백색.
C. ankaranensis - Flowers, Fruits
- Flowers ⇒ 5-merous (다섯 갈래);
- pedicel (꽃자루) ⇒ 길이 3-7(-8) ㎜, 포엽(bractlets) 1개 또는 2개도 있음;
- bractlets (포엽) ⇒ hemi-ellipsoid to ± square, 0.6-1 × 0.5-0.9 ㎜, bearing colleters;
- colleters (분비털) ⇒ 각 포엽에 10-16개, ± cylindrical, 0.4-0.6 ㎜ long, white.
- Calyx (花萼)(hypanthium)
⇒ ± 팽이모양(turbinate), 1.4-1.7 × 2.4-2.7 ㎜, 약간 거친 질감,
⇒ 매우 드문드문 돌기가 있음(papillate) 또는 성기게 매우 짧고 미세한 가시들(echinate)로 덮여 있음; - calyx limb (꽃받침 테) ⇒ truncate (截形), 디스크 보다 훨씬 더 짧음.
- Corolla (꽃부리, 花冠) ⇒ 13.5-17 × 12-14 ㎜,
- corolla tube (꽃부리통, 花冠筒) ⇒ ± corolla lobes와 같거나 약간 더 짧음, 매끄러움;
- corolla tube ⇒ ± 매우 넓게 깔대기 모양(funnel-shaped), 길이 7-9 ㎜ long;
- corolla lobes (花冠裂片) ⇒ 7-8 × 3.6-4 ㎜.
- Stamens (수술, 雄蘂):
- Filaments (수술대, 花絲) ⇒ 下部側着 (infradorsifixed), 길이 2.5-2.7 ㎜;
- anthers (꽃밥, 花葯) ⇒ 좁게 타원형~선형, 길이 5.7-6.3 ㎜.
- Ovary (씨방, 子房):
- disc ⇒ 현저, ± 圓盤모양(discoid)-管形(tubular).
- Style (암술) ⇒ 길이 11.5-14.5 ㎜;
- stigma lobes (암술머리 편) ⇒ 길이 2.3-2.6 ㎜.
- Fruit (immature),
⇒ 타원형~타원-도란형, 2.3-2.5 × 1.1-1.3 ㎝, 녹색; - fruit wall ⇒ 두께 1.5-2 ㎜;
- pedicel (꼭지) ⇒ 길이 7-9 ㎜ long;
- calyx limb (꽃받침 테) ⇒ 눈에 잘 안 띔(inconspicuous).
- Seeds (immature)
⇒ 타원형~타원-도란형, c. 15 × 7-8.7 ㎜, c. 9 ㎜ thick, brown (when dry). - —Fig. 1.
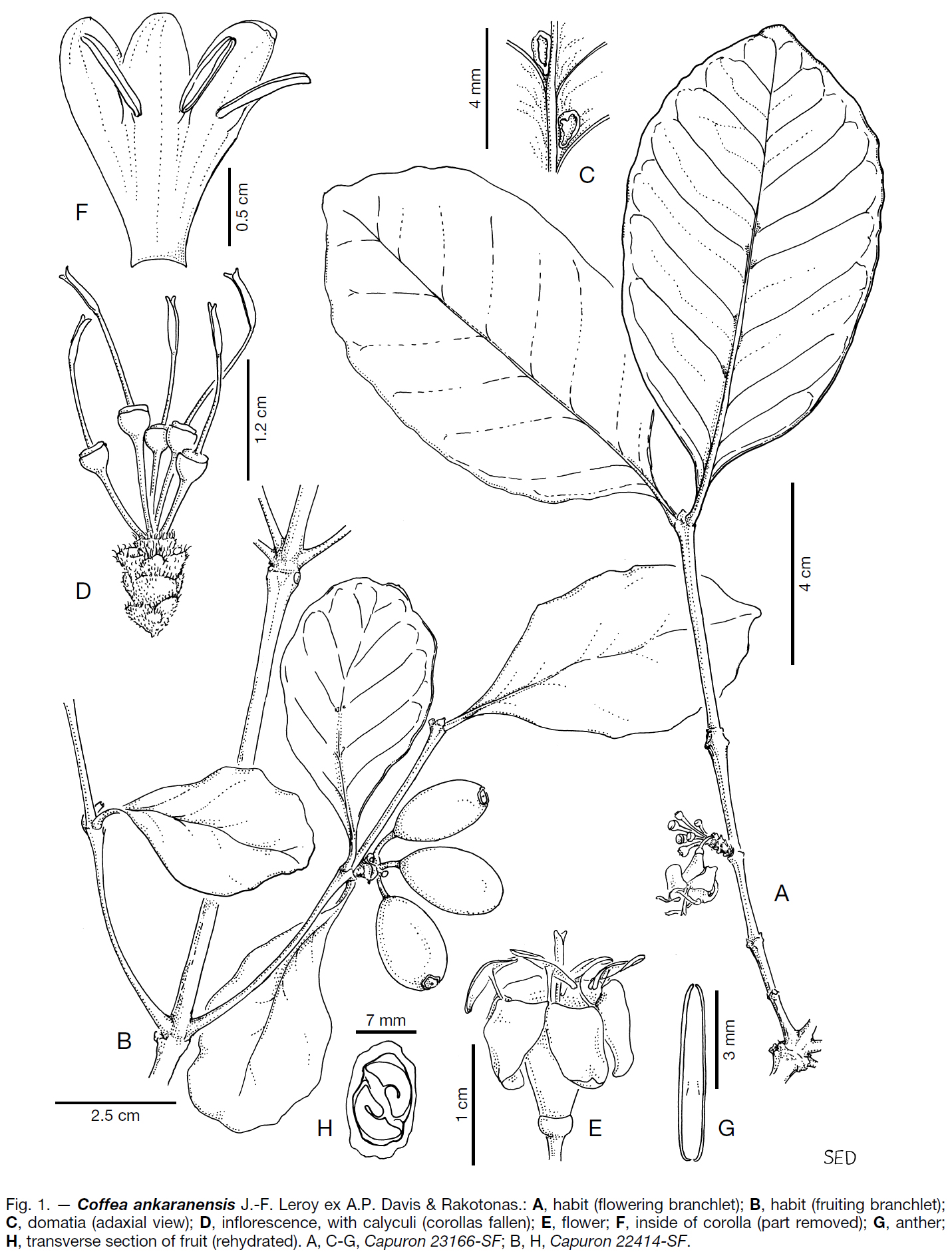

C. ankaranensis - DISTRIBUTION (分布). —
- 마다가스카르 북부, 안치라나나 지방(Antsiranana Province);
- 대개 Ankarana Massif와 근처 숲에 제한적이지만, Forest of Sakaramy에서 나온 2가지 컬렉션들이 있기는 함 (due north east of Montagne d’Ambre, c. 12º26’S, 49º16’E).
- —Fig. 2.
C. ankaranensis - HABITAT AND ECOLOGY (棲息地 및 生態). —
- 계절적으로 건조한 숲: 낙엽수림 또는 낙엽수-상록수 혼합림.
- 석회암(limestone) 및 현무암(basalt lavas).
- 고도 200~600 m.
C. ankaranensis - PHENOLOGY (生物季節學). —
- 11월과 12월에 개화; 1월과 2월에 결실.
- 재배된 표본의 계절 자료는 포함되지 않았음.
C. ankaranensis - CONSERVATION STATUS (保存 狀態). —
- IUCN Red List Category: Endangered (EN B1 a,b).
B1 — total extent of occurrence less than 5000 ㎢ (c. 500 ㎢);
a. — severely fragmented, and possibly existing at no more than five locations;
b (i-v). — continuing decline, observed and inferred. - 사카라미(Sakaramy) 숲에서의 필드워크 (DAVIS & RAKOTONASOLO, 개인 관찰, 1988)는 C. ankaranensis를 발견하지 못했다; 이 숲은 현재 크게 줄어들고 심하게 훼손되었다.
이 증거는 잘못 기록되었거나(식물 표본 지역 데이터가 부정확함) 현재 이 지역에서 멸종되었음을 시사한다.
사카라미에서 C. ankaranensis가 발견되지 않으면 출현 범위와 점유 면적이 상당히 줄어들어 이 種이 Critically Endangered Category (CR)에 속하게 될 가능성이 있다. - 일부 C. ankaranensis 개체군은 보호 구역, 즉 Réserve Spéciale d’Ankarana에 위치해 있다.
C. ankaranensis - PARATYPES. —
- N MADAGASCAR, Antsiranana:
- Capuron 28723-SF, Ouest (Nord): lapiaz dans les calcaires du Mur de l’Ankarana, 16-28 Jan. 1969, fr. (P);
- Guillaumet 2318, face Ouest de l’Ankarana de Diego-Suarez, village d’Andrafiabe, 19 Jan. 1969, fr. (P, TAN);
- Capuron 24554-SF, Ouest (Nord): plateau de l’Ankarana: près d’Ambondromifehy, 5 Feb. 1966, fr. (K, P, TAN, TEF);
- Humbert 32397, forêt de Marovato, canton d’Anivorano Nord, 30 Jan. 1960, fr. (K, P);
- Vianney A. 529 (herb. Leroy 3-43), forêt d’Antsandoko, 9 Nov. 1966, ster. (P);
- Davis & Rakotonasolo 2331 (Kianjavato acc. no. A. 525), Andranofehy, recollected from Coffee Research Station at Kianjavato (FOFIFA), recollected 27 Nov. 1999, fl. (K, P, MO, TAN);
- coll. ignot. A. 526 (herb. Leroy 3-38), forêt d’Antsandoko, 8 Nov. 1966, ster. (P);
- Guillaumet 2187, route de Jofferville, km 4.5 (Diego-Suarez) [locality not traced], 200 m, 4 Aug. 1968, ster. (P, TAN);
- coll. ignot. A. 525 (herb. Leroy 3-36), forêt d’Antsandoko, Ankarana, 8 Nov. 1966, ster. (P, TAN);
- coll. ignot. A. 518, forêt de Sakaramy, 6 Nov. 1966, ster. (K, P);
- Davis & Rakotonasolo 2336 (Kianjavato acc. no. A. 518), forêt de Sakaramy, recollected from the Coffee Research Station at Kianjavato (FOFIFA), recollected 28 Nov. 1999, fl., fr. (K, P, MO, TAN);
- coll. ignot., s.n., forêt de Sakaramy, 13 Nov. 1970, fl. (P).
C. ankaranensis - vs. C. perrieri, Etymology
- 코페아 앤카라넨시스(Coffea ankaranensis)는 다른 Coffea 종에서는 볼 수 없는 특성의 조합이 특징이다.
- 주요 특성은 다음과 같다.
잎은 타원형에서 도란형 또는 넓은 타원형이며, 일반적으로 너비에 비해 다소 넓다(보통 5.4-12 × 4.4-6.2 cm).
외악은 솜털(pubescent)에서 융모(villous)이며, 꽃(13.5-17 × 12-14 ㎜)과 열매는 다소 크다 (2.3-2.5 × 1.1-1.3 cm). - Coffea ankaranensis는 C. perrieri Drake ex Jum. & H. Perrier와 유사하지만, 이 종의 잎은 타원형에서 타원-장방형(elliptic-oblong), 좁은 타원형 또는 타원-倒矛形(oblanceolate, 거꿀창모양)이며 너비에 비해 특별히 넓지 않다 (보통 9.5-13 × 3-5.5 cm); 외악은 유모가 있고(puberulous), 꽃(10-13 × 6-14㎜)과 열매는 더 작다(1.1-1.2 × 0.9-1 cm).
- 이 두 종의 생태는 상당히 다르다.
- Coffea ankaranensis는 일반적으로 석회암과 현무암 위의 계절적으로 건조한 낙엽수림에서 발견되는 반면,
- C. perrieri는 거의 항상 강가 모래(riverine sand) 또는 硅巖(quartzites) 위의 갤러리 숲(gallery forest)으로 제한된다.
- Coffea ankaranesis는 마다가스카르 북부의 Antsiranana 지방에 있는 큰 석회암 大山塊인 앤카래나(Ankarana)에서 유래했기 때문에 그렇게 명명되었다.
Coffea sambavensis
Coffea sambavensis J.-F. Leroy ex A.P. Davis & Rakotonas., sp. nov.
Coffeae millotii J.-F. Leroy affinis sed ramulis apicem versus albidis (nec brunneis), venis lateralibus (venis secundariis) foliorum 6-8-jugis [nec (6-)8-10-jugis], lamina in superficie abaxiali laevi (nec rugosicoriacea), domatiis in superficie abaxiali prominentibus (nec obscuris) etiam in superficie adaxiali manifestis (nec invisibilibus), a costa distantibus 2.5-8 mm (nec juxta costam) positis, distinguenda.
TYPUS. — Capuron 27706-SF, Madagascar, Est (Nord): forêt littorale, au Sud de Sambava, 1-10 Apr. 1967, fr. (holo-, P!; iso-, K!, TEF!).
C. sambavensis - Small tree
- Small tree, ⇒ 樹高 4-8 m high, 胸高 直徑 dbh 4-16 ㎝.
- Bark ⇒ whitish to grey, slightly rough to rough and peeling in patches c. 7 ㎝ in diam.
- Branches
⇒ terete to 4-angled, 直徑 5-8 ㎜, grey to whitish, or brown,
⇒ smooth, sometimes soft and peeling. - Branchlets
⇒ terete to 4-angled, 直徑 2.5-5 ㎜, white to brown,
⇒ usually soft and spongy with freely peeling bark,
⇒ 가끔 많은 작은 검은 반점들이 있음 (?lenticels (皮目) or ?asci (子囊)). - Stipules (턱잎, 托葉)
⇒ caducous (일찍 떨어짐), 卵形(ovate)~넓은 卵形 또는 ± 델타형, 2.2-3 × 3.5-4.2 ㎜,
⇒ 약간 가죽 같음, 무모, 둘레는 無毛;
⇒ apex → 鈍頭(obtuse)~銳頭(acute).
C. sambavensis - Leaves
- petioles (잎자루, 葉柄) ⇒ 0.9-1.5 ㎝ long;
- leaf-blades (잎새, 葉片)
⇒ 넓게 卵形(ovate)~卵形-둥근형(orbicular), 도란-둥근형, 넓게 倒卵形, 또는 길쭉- 倒卵形,
⇒ (5.2-)8-9.7(-10.5) × (2.2-)3.5-7.7 ㎝, 약간 가죽 같은; - Base (잎밑, 葉底) ⇒ 流底(attenuate);
- Margins (잎 둘레, 葉緣) ⇒ 약간 反曲~反曲(revolute);
- apex (잎끝, 葉先) ⇒ 原頭~鈍頭, 종종 약간 漸尖頭(subacuminate);
- abaxial surface (背軸面, 下面):
- Midrib (主脈) ⇒ 顯著(prominent);
- Secondary veins (2차 맥)
⇒ 현저, 6-8 쌍, 30°~45° 각도로 올라감, 직선~곡선,
⇒ 결합하여 고리형 둘레맥을 형성, 둘레와 더 가까운 2차 둘레맥도 있음; - tertiary venation (3차 맥) ⇒ 뚜렷~현저, ± 網狀;
- higher order venation (그 이상 맥) ⇒ 모호(obscure), 分岐됨(ramified);
- adaxial surface (向軸面, 上面):
- 脈 ⇒ 배축면의 경우보다 더 명확하게 뚜렷함;
- domatia (소혈)
⇒ 움 타입(crypt type), 몇 개 안되지만 눈에 잘 띔(conspicuous),
⇒ 보통 2차 맥을 따라 주맥에서 2.5-8 ㎜에 위치,
⇒ 구경 0.1-0.3 ㎜, 또는 괴사되어가는 소혈들이 있으며 구경은 0.4-10 ㎜,
⇒ 둘레는 약간 올라옴 또는 부풀어 오름, 그리고 종이 같음(chartaceous), 무모;
⇒ 향축면에서 현저, 작은 타원형 농포처럼 뚜렷, 1.5-3.5 × 1-1.2 ㎜, 가끔 잎 면보다 더 진함.
C. sambavensis - Inflorescence, Calyculi
- Inflorescence (꽃차례, 花序)
⇒ 엽액당 1개, 2-4(-6)개 꽃이 핌,
⇒ 매우 짧게 가지 발생, 기저부위에 1개의 짧은 가지 (0.2-0.3 ㎜ long)가 있음,
⇒ 길이 5-6 ㎜, 열매 발달 중에 상당히 신장,
⇒ 종종 삼출액으로 약간 덮여 있음; - 花序軸 inflorescence axis (bearing calyculi) ⇒ 길이 4.5-6.5 ㎜.
- Calyculi (外萼)
⇒ 3개, 기저 및 낮은 외악들이 종종 떨어지거나 부서짐,
⇒ ± sessile (착생), 약간 가죽 같음, 무모~柔毛,
⇒ 둘레 → 무모 또는 적은 수의 털 (0.2-0.3 ㎜ long); - basal (1st) calyculus
⇒ 1.8-2.2 × 2.4-3.6 ㎜,
⇒ stipular lobes 1.7-2 × 1.6-2.7 ㎜,
⇒ foliar lobes not seen; - middle and upper (2nd and 3rd) calyculi
⇒ (2.5-)4.3-5.5 × (3.2-)4.9-12 ㎜,
⇒ stipular lobes broadly ovate to ± deltate, (2-)3-5.2 × 3-5 ㎜,
⇒ foliar lobes elliptic to obovate, distinctly smaller than stipular lobes, 1-1.5 × 0.3-0.5 ㎜; - internal surfaces of calyculi
내면 ⇒ 많은 분비털들이 있음, 특히 기저부분에, 많은 가는 털들이 섞여 있음 (0.2-0.3 ㎜ long);
colleters (분비털) ⇒ 좁게 원추형~매우 좁게 타원형, 길이 약 0.2 ㎜, 백색.
C. sambavensis - Flowers, Fruits
- Flowers
⇒ 다섯 갈래 (5-merous);
⇒ 꽃자루 길이 2-5.5 ㎜, 포엽(bractlets) 없음. - Calyx (꽃받침, 花萼) (hypanthium, 꽃턱통, 花托筒)
⇒ ± 거꿀원뿔형(obconical)~鐘 모양(campanulate), 1.8-2.3 × 1.8-2.4 ㎜, glabrous, smooth;
⇒ calyx limb → 매우 좁게 5-편~截形(truncate), 디스크 보다 더 짧음;
⇒ disc → 다소 현저, ± discoid. - Corolla (꽃부리, 花冠) ⇒ 15-17 × 11-14 ㎜, corolla lobes 보다 corolla tube이 약간 더 김, 매끄러움;
- corolla tube (花冠筒) ⇒ ± 넓게 깔대기 모양(funnel-shaped), 길이 9-11 ㎜;
- corolla lobes (花冠裂片) ⇒ 6-8 × 4.7-6 ㎜.
- Stamens (수술, 雄蘂) :
- filaments (수술대, 花絲) ⇒ 下部側着 (infradorsifixed), 길이 4-4.5 ㎜;
- Anthers (꽃밥, 花葯) ⇒ 좁게 타원형~선형, 길이 약 6 ㎜.
- Style (암술) ⇒ 길이 15-16 ㎜;
- stigma lobes (암술머리 편) ⇒ 길이 2.5-3 ㎜.
- Fruit (immature, 덜 익은 것),
⇒ 짧게 도란형~ ± 팽이모양(turbinate), 17-22 × 14-18 ㎜,
⇒ 약간 二葉的(slightly bilobed)~二葉的(bilobed) (upon drying),
⇒ 마르면 상당히 오그라듦(shrinking), 녹색;
⇒ Fruit wall → 두께 3.5-4 ㎜;
⇒ Pedicel (꼭지) → 길이 (6-)9-13 ㎜;
⇒ Calyx limb (꽃받침 테) → 눈에 잘 안 띔(inconspicuous). - Seeds (immature)
⇒ ± 타원형체~타원-도란형체,
⇒ 14-16 × 9-12 ㎜, 두께 약 7 ㎜,
⇒ white (fresh) ~ 연갈색 (when dry). - —Fig. 3.

C. sambavensis - DISTRIBUTION (分布) .—
- 마다가스카르 북동부, 안치라나나(Antsiranana) 지방에 토착적;
- Sambava와 Vohimarina 사이 (Vohémar).
- —Fig. 2.
C. sambavensis - HABITAT & ECOLOGY (棲息地 및 生態). —
- 저지대 다습 상록림, 연안지대(littoral zone) 포함.
- 散砂(loose sands), 基盤岩(basement rocks) 그리고 용암(lavas).
- 고도 0~200 m
C. sambavensis - PHENOLOGY (生物季節學). —
- 개화 11월; 결실은 아마도 4월.
- 재배되는 표본의 계절은 포함되지 않았음.
C. sambavensis - CONSERVATION STATUS (保存 狀態). —
- IUCN Red List Category: Endangered (EN B1 a,b).
- B1 — total extent of occurrence less than 1000 ㎢ (c. 350 ㎢);
- a. — severely fragmented, and possibly existing at no more than five locations;
- b(i-v). — continuing decline inferred.
- 현행 보호구역 체계 내에는 C. sambavensis 개체군이 없다.
- 멸종 위험을 보다 확실하게 평가하려면 Sambava 근처의 남아 있는 1차림(열화된 1차림 포함) 지역과 마다가스카르 북동부 해안을 따라 현장 조사가 필요하다.
C. sambavensis - PARATYPES. —
- NE MADAGASCAR, Antsiranana:
- Capuron 24928-SF, district Est (Nord): forêt littorale, au Sud de Sambava, 20 Oct. 1966, fl. bud (K, P, TEF);
- Davis & Rakotonasolo 2346 (Kianjavato acc. no. 955), Sambava, recollected from the Coffee Research Station at Kianjavato (FOFIFA), recollected 28 Nov. 1999, fl. (K, P, TAN);
- Davis & Rakotonasolo 2323 (Kianjavato acc. no. A. 950), ibid., recollected 27 Nov. 1999, fl., fr. (BR, K, P, MO, TEF);
- Rakotonasolo RNF 270, Vohémar, Fanambana, 190 m, 24 Nov. 2000, fl. bud. (K, P, MO, TAN, TEF);
- Rakotonasolo RNF 274, Sambava, Anjangoveratra, Ambodisambalahy, 26 Nov. 2000, fl. (K, TAN, TEF).
C. sambavensis - vs. Coffea millotii, Etymology
- 코페아 샘바벤시스(Coffea sambavensis)는 다음과 같은 특징의 조합으로 인해 쉽게 알아볼 수 있는 종이다.
가지는 보통 희끄무레하고, 보통 부드럽고 벗겨지는 나무껍질이 있다.
잎은 넓고(길이 대 너비 비율 3:2 이상), 약간 가죽 같은 질감이며, 현저한 도마티아가 있다.
도마티아는 주맥에 붙어 있지 않고, 2차 맥을 따라 몇 밀리미터 떨어진 곳에 있으며, 종종 향축면에서 볼 수 있다 (Fig. 3A, B 참조).
과실은 다소 크고, 벽이 두꺼우며(대부분 중과피), 긴 꽃자루에서 열린다.
꽃자루는 꽃에서는 짧지만, 과실이 자라는 동안 상당히 길어진다.
모든 측정값은 기술을 참조. - Coffea sambavensis는 마다가스카르 동부의 다습림에서 나온 또 다른 종인 코페아 밀로티 (C. millotii J.-F. Leroy)와 가장 유사하다.
- 코페아 밀로티 (Coffea millotii)는 과실과 꽃차례 형태가 C. sambavensis와 유사하지만,
domatia가 주맥에 위치하고
각 잎의 배축면(abaxial surface)이 거칠고 주름져 있기 때문에 (C. sambavensis는 매끄러움) 쉽게 구별할 수 있다.
잔가지 껍질의 색상과 2차 맥의 쌍 수는 위의 진단에서 주어진 것처럼 이러한 종을 분리하는 데 사용할 수 있는 다른 특징이다. - C. millotii의 분포는, 그 분포의 북쪽 한계가 Sambava에서 남쪽으로 약간 떨어진 Masoala 반도에 있기 때문에 C. sambavensis와 겹치지 않는다.
REFERENCES
- BRIDSON D.M. 1988. — Coffea, Rubiaceae: 415-474, in POLHILL R.M., BRIDSON D.M. & VERDCOURT B. (eds), Flora of Tropical East Africa, part 2. Balkema, Rotterdam/Brookfield.
- BRIDSON D.M. 1994. — Additional notes on Coffea (Rubiaceae) from tropical East Africa. Kew Bull. 49: 331-342.
- CHEVALIER A. 1942. — Caféiers du globe, fasc. 2 : iconographie des caféiers sauvages et cultivés. Encycl. Biol. 22: 1-36, 158 plates.
- CHEVALIER A. 1947.—Caféiers du globe, fasc. 3 : systématique des caféiers et faux-caféiers maladies et insectes nuisibles. Encycl. Biol. 28: 1-352.
- DAVIS A.P. & RAKOTONASOLO F. 2000.—Three new species of Coffea L. (Rubiaceae) from Madagascar. Kew Bull. 55: 405-416.
- DAVIS A.P. 2001. — Two new species of Coffea L. (Rubiaceae) from eastern Madagascar. Kew Bull. 55: 479-489.
- DAVIS A.P. & RAKOTONASOLO F. 2001.—Three new species of Coffea L. (Rubiaceae) from NE Madagascar. Adansonia, sér. 3, 23: 137-146.
- HUMBERT H. 1955. — Les Territoires Phytogéographiques de Madagascar. Leur Cartographie. Colloque sur les Régions Ecologiques du Globe, Paris, 1954. Année Biol. 31: 195-204.
- IUCN 2001. — IUCN Red List Categories, Version 3.1., prepared by the IUCN Species Survival Commission. Gland, Switzerland and Cambridge.
- LEROY J.-F. 1961.—Coffeae novae Madagascarienses. J. Agric. Trop. Bot. Appl. 8: 1-20.
- LEROY J.-F. 1962a. — Prospection des Caféiers sauvages : rapport préliminaire sur une mission scientifique à Madagascar et aux Iles Mascareignes. J. Agric. Trop. Bot. Appl. 9: 211-244.
- LEROY J.-F. 1962b.—Coffeae novae Madagascariensis et Mauritianae. J. Agric. Trop. Bot. Appl. 9: 525-530.
- LEROY J.-F. 1972a. — Prospections des caféiers sauvages de Madagascar : deux espèces remarquables. Adansonia, n.s., 12: 317-328.
- LEROY J.-F. 1972b. — Prospections des caféiers sauvages de Madagascar : sur deux espèces sympatriques du Nord. Adansonia, sér. 2, 12: 345-358.
- ROBBRECHT E. 1978.—Sericanthe, a new African genus of Rubiaceae (Coffeeae). Bull. Jard. Bot. Nat. Belg. 48: 3-78.
- STOFFELEN P. — 1998. Coffea and Psilanthus in tropical Africa: a systematic and palynological study, including a revision of the West and Central African species. PhD Thesis: Katholieke Universiteit Leuven.
'Coffee Botany' 카테고리의 다른 글
| 꼭두서니科 코페아屬의 분류학적 개요 (A.P. Davis et al. 2006) (0) | 2024.10.31 |
|---|---|
| 코페아 마피아나 – 카메룬 南部의 왜소성 코페아 新種 (2) | 2024.10.19 |
| 코페아 리토랄리스, 코페아 맥퍼슨니, 코페아 라치마망게 (0) | 2024.10.13 |
| 코페아 보헤마렌시스, 코페아 미뉴티플로라, 코페아 모라티 (1) | 2024.10.01 |
| 코페아 라코토나소로이, 코페아 몬티스-새크리 (0) | 2024.09.29 |



