1세대 DNA 시퀸싱 (염기서열분석) 포스트에서 언급된 바와 같이, 시험관 내 증폭(in vitro amplification) 및 핵산 검출 (detection of nucleic acids), 병렬 처리(parallel processing) 및 생물정보학(bioinformatics)과 같은 다양한 분야의 기술적 혁신 (Diaz et al., 2011, 2014; Esteban et al., 2013; Galvez et al., 2010, 2011)은 당시 ‘next-generation 차세대’라고도 알려진 2세대 핵산 서열 분석 플랫폼의 개발을 가능케 했다. 여기에는 대량의 스타팅 핵산 분자가 필요한 전략이 포함되어 최신 기술과 비교할 때 중간 처리량 결과를 생성한다.
2005년부터 이러한 새로운 어푸로치들을 통해 단일 기계 실험에서의 시퀀싱 생산성을 수 kilobases (kb)에서 Gigabases (Gb)로 높일 수 있었다. 이는 6 자릿수 이상 (즉, 100만 배 증가)를 나타낸다. 5가지의 2세대 핵산 시퀀싱 전략들이 현재까지 공개되었다 (이 중 4개는 상용 플랫폼 장비임).
| 1970s | 1990s | 2000s | 2010s | 2020s | |
| 세대 | 겔-기반 방법들의 1세대 |
모세관 방법들의 1세대 |
차세대 | 3세대 | 미래 세대 |
| 기술 | Sanger sequencing (manual) | Sanger sequencing (automated) | Illumina, Roche 454, Ion Torrent | PacBio SMRT, ONT | Genapsys, MGI |
| 특징 | 겔-기반 분석 | 모세관 분석 | 높은 결과물 | 롱 리드 | |
| 찬성 | 정확성 | 더 높은 규모, 더 낮은 비용, 정확성 | 롱 리드, 울트라-패스트, 수익성 | 비용 효과성 목표, 정확성 |
|
| 비판 | 노동 집약적, 고 비용 |
숏 리드가 분석을 어렵게 함 | 정확성 감소 | ||

로쉬 454 DNA 시퀀싱 (Roche 454 Life Sciences Emulsion-PCR) 기술은 가장 긴 리드(reads)을 생성하는 2세대 시퀀싱 방법이다 (200에서 1000b까지)(Margulies et al., 2005). 454 시퀀싱은 2005년 Roche社가 개발한 최초의 성공적인 차세대 시퀀싱 방법이었다. 이는 32억 개의 염기서열을 분석하는 데 13년이 걸린 인간 게놈 프로젝트의 일부로 사용된 이전 기술보다 훨씬 빨랐다. 이에 비해 454 시퀀싱은 하루에 최대 10억 개의 염기를 시퀀싱할 수 있다. 그러나 플랫폼에는 약점이 있었고 더 정확한 방법을 선호하여 2010년대 중반에 중단되었다.
로쉬 454 시퀀싱 절차 요약
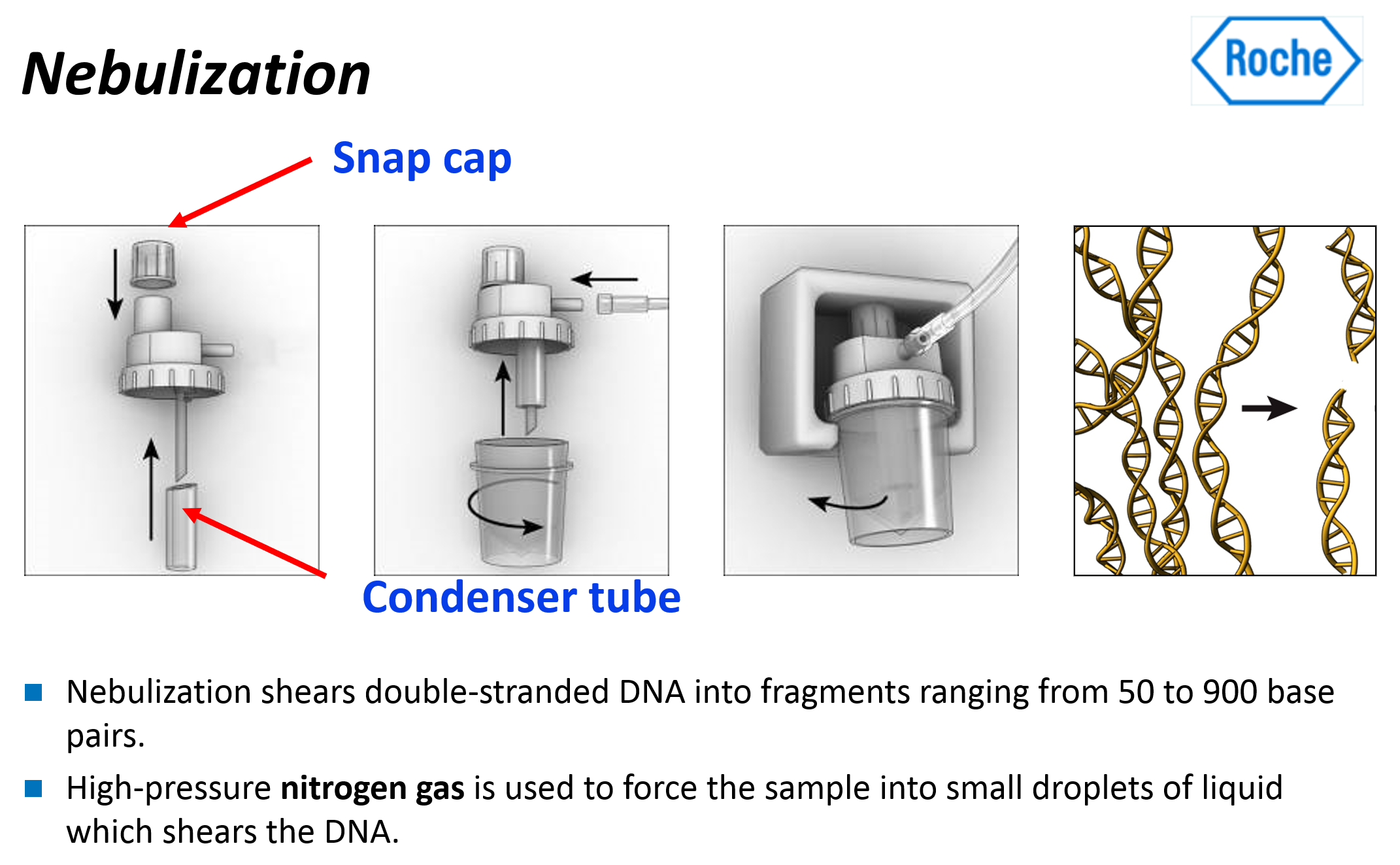
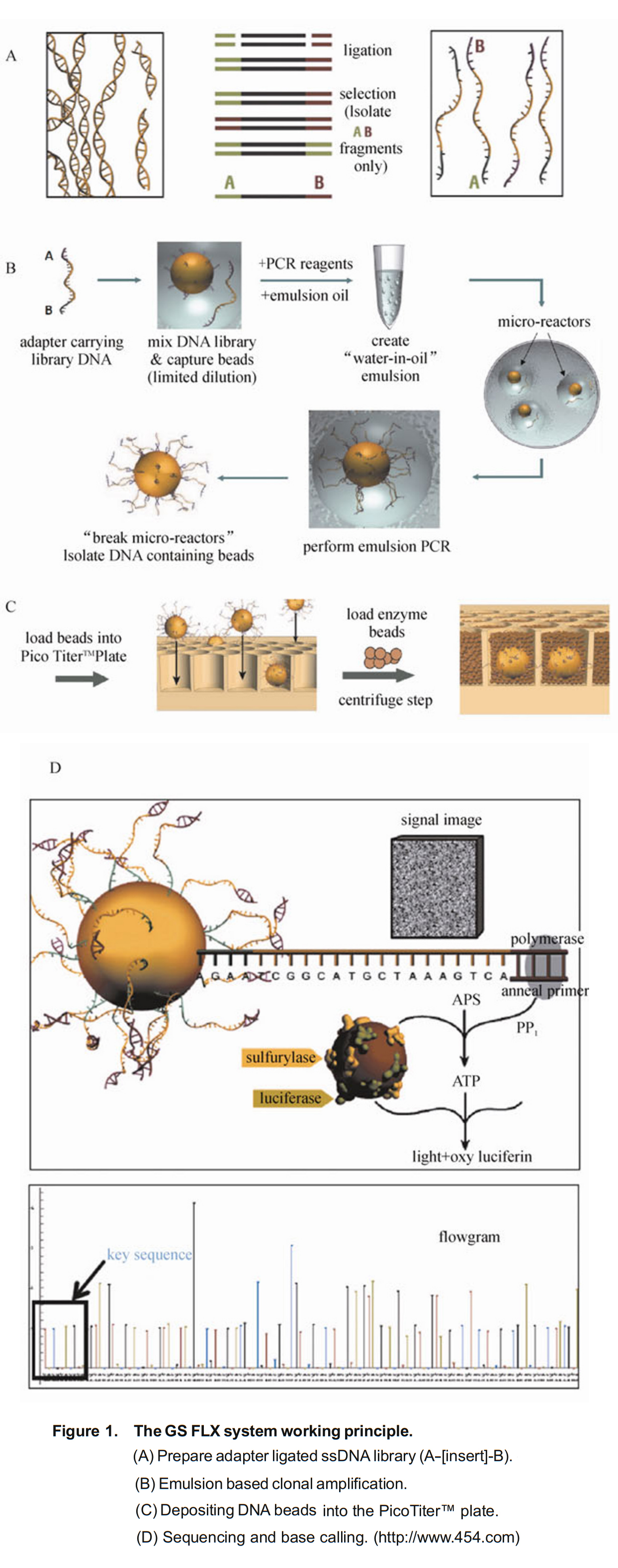
- 분무(nebulization)를 통해 dsDNA의 무작위 단편(random fragments)을 생성하는 과정이 포함된다.
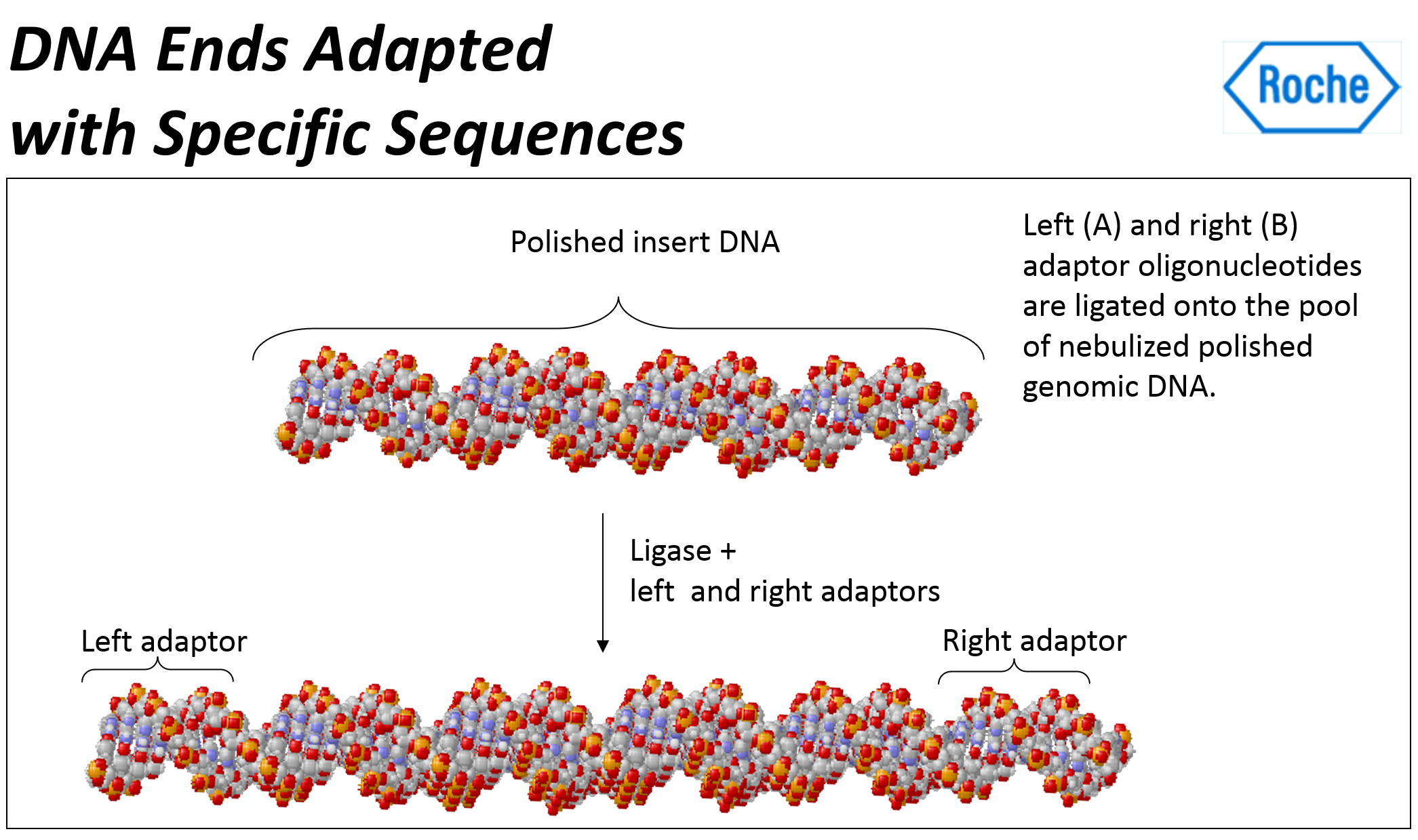
- 그런 다음 이러한 어댑터 중 하나에 있는 비오틴(biotin)을 포함하여 특수 어댑터(Special adapters)를 조각 끝 부분(fragment ends)에 결찰한다 (ligated).
- 이를 통해 dsDNA를 스트렙타비딘 코팅 비드 (streptavidin-coated beads)에 고정할 수 있으므로, 변성 (denaturation) 후 ssDNA 가닥 (strands)을 효과적으로 분리할 수 있다.
- 싱글 분리된(single isolated) ssDNA 가닥은 프라이머 코팅된 비드(primer-coated beads)에 의해 포획되며 (captured),
- 이는 유중수 에멀젼 (water-in-oil emulsion)의 작은 PCR 반응기에서 분리된다.
- 포획된 ssDNA는 비오티닐화된 프라이머 (biotinylated primers)를 사용하는 emulsion PCR (emPCR)에 의해 증폭된다.
- 그런 다음 에멀젼을 깨뜨리고 스트렙타비딘 코팅 비드를 사용하여 DNA를 포획하고 DNA를 변성시킨다.
- 포획된 ssDNA는 분리되어 피코웰(picowells)에 배치되며, 여기에서 파이로시퀀싱 반응 (pyrosequencing reaction)이 수행되어 각 dNTP를 하나씩 플러싱 (flushing)한다.
- DNA 중합효소(polymerase)에 의해 뉴클레오티드가 추가되면, 설퍼릴라제 (sulfurylase), 루시퍼라제 (luciferase) 그리고 루시페린 (luciferin)과 관련된 반응이 일어나 빛(光, light)을 생성하며, 이 빛은 수십만 개의 피코웰(picowells) 각각에 대해 동시에 검출기에 의해 기록된다.
- 그런 다음 생물정보학 도구를 사용하여 DNA reads를 콘티그(contigs)로 조립하고, 이에 따라 염색체와 게놈이 생성된다 (Figure 3).

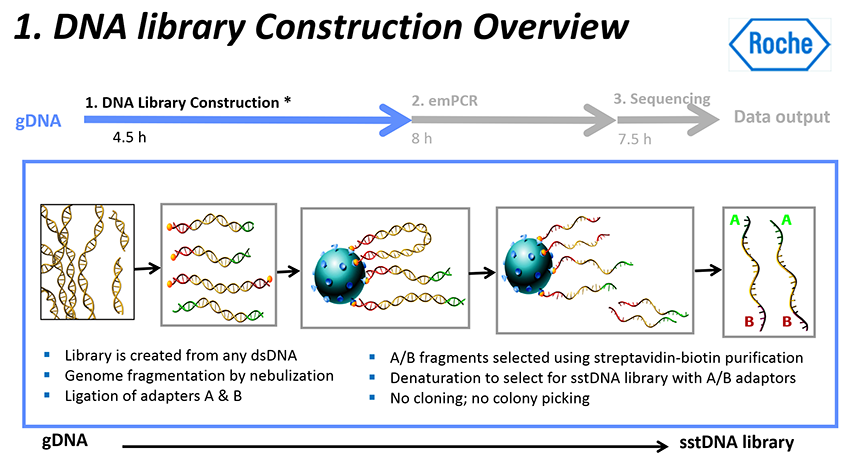
로쉬 454 시퀀싱 절차 : DNA Library Construction
- 이 과정은 이중 가닥(double-stranded) DNA (dsDNA) 샘플로 시작된다.
- Spray method 또는 분무(nebulization)를 사용하여 DNA 샘플들을 300~800개의 작은 fragments로 분해한다.
- 양쪽 끝에 어댑터들(adapters)을 더해준다.
즉, 어댑터라고 불리는 짧은 DNA 서열이 DNA 단편에 부착된다. 양쪽 끝에 다른 어댑터들을 부착한다. - "A"어댑터와 "B"어댑터는 길이가 44 bases이고, 그중 20 bases는 PCR 프라이머 요소이고, 20 bases는 시퀀싱 프라이머이며, 4개는 key이다. "B"어댑터에만 5' end에 Biotin이 포함되어 있다.
- 어댑터들은 한쪽 끝에서는 뭉툭하고, 다른 한쪽 끝은 오목하여 단편들에 결찰되도록 되어 있다.
- 어댑터 결찰 결과, dsDNA 양쪽에 어떤 어댑터가 붙었는지에 따라, 4가지 타입들이 나온다 (AB, AA, BB, BA).
- 이들에 작은 수지 비드(resin beads) (마그네틱 입자) 스트렙타비딘 코팅 비드 (streptavidin-coated beads)를 섞어주면, AB와 BB가닥들이 마그네틱 입자 비드에 직접 결합한다. 비오틴이 없는 AA가닥들은 결합되지 않는다.
- DNA 단편은 비드에 직접 결합하며, 이상적으로는 각 비드에 하나의 단편이 결합된다.
- DNA 단편이 비드의 DNA에 부착되면 이중 가닥을 연결하는 결합이 끊어지고 가닥이 분리되어
단일 가닥 DNA(single stranded DNA, ssDNA)가 된다.
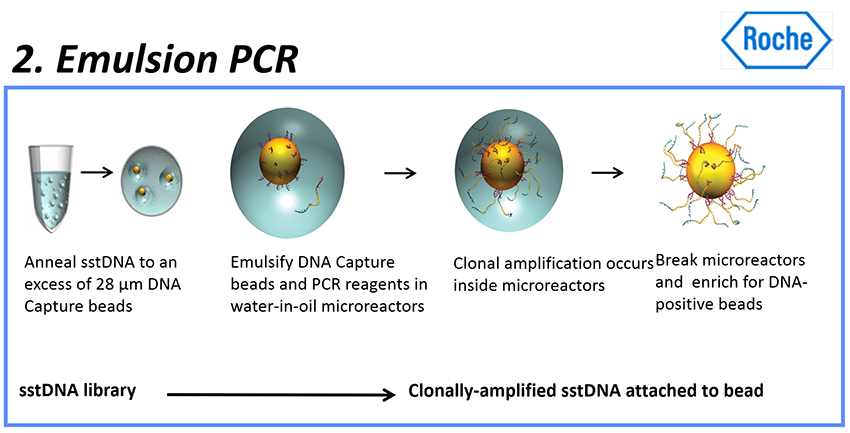
로쉬 454 시퀀싱 절차 : emPCR
- 그런 다음 DNA 단편들은 중합효소 연쇄 반응(polymerase chain reaction, PCR)으로 알려진 반응에 의해 각 비드에 여러 번 복제된다.
- emulsion에 들어있는 28μm 비드들에 의해 고정될 것이다.
- emulsion PCR (emPCR)의 핵심 기술은 에멀젼의 특징을 사용하여 서로 다른 비드들을 분리하는 것이다.
- 샘플 DNA 증폭 전에, PCR 반응의 모든 구성요소들을 담은 수용액이 고속으로 회전하는 미네랄 오일의 표면으로 주입되어, 미네랄 오일로 싸인 수많은 작은 물방울들을 형성한다.
- 하나의 작은 방울이 독립적인 PCR 반응 표면을 형성한다.
- 이상적으로는, 각 작은 물방울이 단 하나의 DNA 템플릿과 하나의 비드만을 가진다.
- 작은 물방울들로 둘러싸인 비드들 표면에는, 그들의 어댑터들과 매치하는 상보적인 올리고들이 있어서, 단일 가닥의 DNA가 그 비드들에 특유적으로 결합할 수 있다.
- 동시에, 인큐베이션 시스템이 PCR 반응제들을 가지고 있어서, 그 비드에 고정된 각 작은 DNA 단편이 증폭을 위한 독특한 템플릿일 수 있다.
- 또한, PCR 산물들은 자성을 가진 비드들과 결합될 수 있다.
- 반응이 이뤄진 후에, 에멀젼 시스템은 파괴될 수 있고, 타겟 DNA들이 누적될 것이다.
- 각 작은 단편은 약 1백만배 증폭될 것이며, 따라서 시퀀싱 프로세스에 의해 필요한 양적 수준에 달할 수 있게 된다.
- 이는 수백만 개의 동일한 DNA 시퀀스 사본을 생성한다.

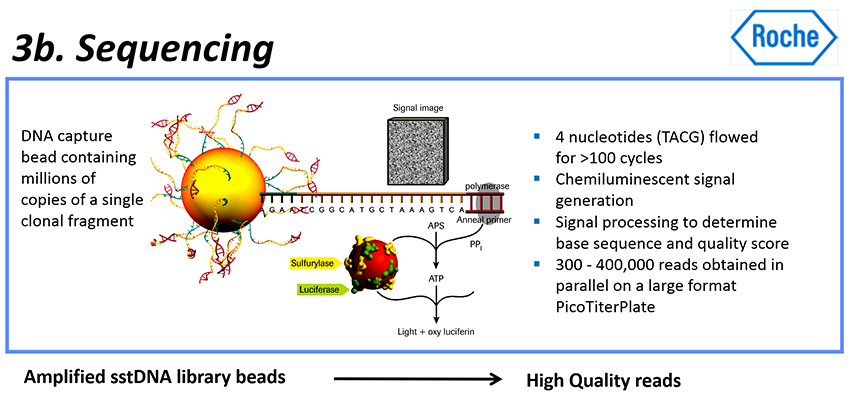
로쉬 454 시퀀싱 절차 : Pyrosequencing
- 그런 다음 비드들을 필터링하여 DNA에 부착하지 못했거나 두 가지 유형 이상의 DNA 단편을 포함하고 있는 비드들을 제거한다.
- 그런 다음, 이 비드들은 PTP plate에 놓여진다 (아래 Figure 1c).
- 이 플레이트는 직경 44μm의 특수한 나노구멍들을 많이 가지고 있다.
- 그런 다음 나머지 비드들을 시퀀싱 반응에 필요한 DNA 중합효소(polymerase enzyme) 및 프라이머가 포함된 효소 비드들(enzyme beads)과 함께 시퀀싱 플레이트의 웰에 넣는다 (웰당 비드 1개).
- 각 나노구멍들은 한 개의 비드만을 수용할 수 있어서, 이 방법을 통해 각 비드의 위치를 고정할 수 있게 하여 시퀀싱에 편리하도록 해준다.
- 시퀀싱 전에 DNAs를 가진 비드들을 처리하는데 폴리머라제(polymerase)와 단일 가닥 DNA 결합 단백질 (ssDNA binding protein)이 필요하다.
- 중합효소와 프라이머는 비드의 DNA 단편들에 부착된다.
- 이런 시퀀싱 과정에서 사용되는 방법을 pyrosequencing이라고 한다.
- 그 나노 구멍으로 작은 비드를 넣고, 시퀀싱 반응을 시작한다.
- DNA 시퀀싱 반응은 증폭되어 고정된 단일 가닥 DNAs에 기초한다.
- 뉴클레오티드 염기들은 한 번에 한 가지 타입의 염기 파동(waves)(As의 웨이브, 이어서 Cs의 웨이브, 이어서 Gs, 그 다음에 Ts)으로 웰에 더해진다.
- 각 염기가 DNA에 결합되면 빛이 나오고 이를 카메라에 기록한다.
빛의 강도는 통합된 같은 타입의 뉴클레오티드들의 수에 해당한다. 예를 들어, 단편들에 3개의 연속적인 A가 있는 경우 생성되는 빛의 양은 조각에 있는 단일 A의 3배이다. - 이러한 빛 강도 패턴을 그래프에 표시하면 오리지널 DNA 조각의 시퀀스를 해독할 수 있다.
- 다른 말로 하면, 하나의 dNTP가 템플릿 DNA와 짝을 지울 수 있으면, 합성 후에 pyrophosphate기가 발생될 것이다.
- 발생되는 pyrophophate기는 ATP sulfuric acid 화학효소들과 반응하여 ATP를 만든다.
- ATP의 CO-oxidation과 luciferase는 fluorescein molecule (플루오레세인 분자, 형광물질 분자)를 촉발시키며 형광을 발하며, 그 PTP 보드의 다른 쪽에 CCD 카메라가 형광 신호를 기록한다.
- 마지막으로, 그 결과는 컴퓨터 소프트웨어에 의해 처리된다.
dNTP의 각 종류는 그 반응에서 독특한 형광색에 따라 측정될 수 있으므로, DNA 시퀀스가 그 형광색에 따라 측정될 수 있다. - 반응 후에는, ATP가 diphosphatase에 의해 분해되어, 플루오레세인을 퀀칭하고, 시퀀싱 반응은 다음 사이클로 간다.
- 454 시퀀싱 기술의 경우에 있어서, 각 반응이 PTP 보드의 독립적인 나노 구멍들에서 수행되므로, 상호 간섭과 시퀀실 바이어스를 크게 줄인다.
- 454 기술의 가장 큰 장점은 긴 시퀀싱 리드 길이를 얻는다는 것이다.
현재 454 기술의 평균 리드 길이는 400bp까지나 된다. - 454는 Illumina의 Solexa와 Hiseq과는 완전히 다른 것이다.
- 454 시퀸싱 기술의 단점은 호모폴리머(homopolymer) 길이를 정확하게 특정할 수 없다는 것이다.
- 이런 피할 수 없는 이유 때문에, 454 기술은 그 결과에 삽입 및 삭제 시퀀싱 에러들을 도입할 것이다.
'Coffee Genetics' 카테고리의 다른 글
| SOLiD Sequencing : 2세대 DNA 염기 서열 분석 (3) | 2024.07.14 |
|---|---|
| Illumina Sequencing Technology - 2세대 DNA 시퀀싱 (22) | 2024.07.11 |
| 1세대 DNA 시퀀싱 (22) | 2024.07.01 |
| 식물 세포, 핵, DNA, 유전자, 게놈 (1) | 2024.06.29 |
| 아라비카 커피의 기원과 多樣化 歷史 (Coffea arabica diversification history) (0) | 2024.06.15 |




댓글