



18.1 Introduction
- 코페아 아라비카(Coffea arabica)는 商業的으로 栽培된 최초의 코페아屬(genus Coffea)의 種(species)이었으며, 따라서 커피 음료의 品質을 評價하는 準據가 되었다.
- 커피 음료의 品質은
재배 過程에서부터 콩의 貯藏, 加工, 로스팅에 이르기까지 다양한 要因, 作用, 그리고 決定의 均衡에 의해 결정된다.
┌ 非遺傳的 要因과
└ 遺傳的 要因 ⇒ 모두 커피 음료의 品質에 影響을 미치는 것으로 나타났다 [1]. - ┌ 土壤 肥沃度(Soil fertility),
├ 施肥(fertilization),
├ 氣候(climate),
├ 高度(altitude),
├ 植物 健康(plant health),
├ 收穫 시 열매의 成熟 段階(stage of fruit maturation),
├ 收穫後 加工(post-harvest processing), 그리고
└ 遺傳的 背景(genetic background)이
⇒ 모두 커피 飮料의 品質에 影響을 미치는 것으로 알려진 主要 要因의 예이다 [1–3]. - 그러나 이러한 모든 要因의 結果는
⇒ 콩들의 最終 化學的 組成(chemical composition)이며,
⇒ 이는 로스팅 過程에서 아로마와 플레이버가 發達하는 동안 劇的으로 변한다. - 커피 化學의 複雜性은
아로마에서 식별되는 1,000개가 넘는 揮發性 化合物의 數에서 가장 잘 드러난다 [4,5]. - ┌ 糖類(Sugars),
├ 아미노산(amino acids),
├ 펩티드(peptides),
├ 脂質(lipids),
├ 트리고넬린(trigonelline),
├ 클로로겐酸(chlorogenic acids),
├ 有機酸(organic acids), 그리고
└ 카로티노이드(carotenoids)가 커피 品質에 影響을 미치는 것으로 알려져 있다 [6,7]. - 또한
┌ 아미노산(amino acids),
├ 펩티드(peptides),
└ 蛋白質(proteins)은
⇒ 로스팅 過程에서 마이야르 反應(the Maillard reaction)을 통해 糖類와 相互作用하기 때문에 [11],
⇒ 커피 아로마 生成에 중요한 役割을 하는 것으로 잘 알려져 있다 [6,8–10].
- 지난 50년 동안 커피콩의 蛋白質 및/또는 아미노산 含量과 프로필에 대한 여러 보고서에서 이러한 化合物이 커피 플레이버에 중요하다는 것이 지속적으로 確認되었다 [9,10,12–14].
- 그러나 이러한 硏究에 사용된 技術과 科學的 接近 方式이 改善되었음에도 불구하고, 이러한 모든 硏究는 相對的 重要性(構成比率 및 濃度)을 명확하게 把握하지 못했다.
- 문헌에는 C. arabica와 C. canephora 콩의 蛋白質 含量이 10–15%로 다양하게 보고되었지만, 이러한 差異는 蛋白質 含量 推定에 사용된 方法에서 비롯된다 [6,15].
- 카페인과 트리고넬린에 대한 보정을 하면 건조 기준으로 약 11.5%의 평균값이 發見되고
C. arabica와 C. canephora 콩 사이에 유의미한 差異는 없는 것으로 보인다 [6,15]. - 總 커피콩 蛋白質의 3분의 1은
⇒ 細胞壁의 아라비노갈락탄(arabinogalactans)에 결합하는 것으로 생각되지만,
⇒ 이러한 蛋白質의 特性에 대한 자세한 정보는 없다. - 대부분이 폴리페놀 酸化酵素(polyphenol oxidase) 및 過酸化酵素(peroxidases)와 같은 酵素(enzymes)일 가능성이 높으며, 蛋白質의 대부분은 細胞質에 남아 있다 [15].
- 遊離 아미노산(free amino acids) 含量은
⇒ C. arabica 및 C. canephora 品種에서 0.15~2.5%로 蛋白質보다 훨씬 낮다 [6,15].
⇒ 그럼에도 불구하고, 遊離 아미노산은 커피 플레이버 發達과 關聯된 가장 중요한 化合物 그룹이다 [6]. - C. canephora 品種은
⇒ C. arabica에 비해 glutamate를 제외한 모든 遊離 아미노산 含量이 더 높다 [6]. - 몇가지 다른 보고서들에서는 커피콩의 酸化還元 狀態(redox status)와 蛋白質 및 커피 品質과의 關係에 대한 探究를 說明했다. 특히 C. arabica의 化合物 중 약 5~7.5%를 차지하는 클로로겐산(chlorogenic acids)에 주목했으며, 다른 Coffea 種의 커피콩에서는 더 높은 水準에 달할 수도 있을 것이다 [6].
- 초기 硏究에서는 폴리페놀 酸化酵素(enzyme polyphenol oxidase)가 이러한 페놀산들(phenolic acids)을 퀴논(quinones)으로 酸化시키고, 이는 다시 蛋白質과 反應하여 최종 콩 品質과 결과적으로 飮料에 影響을 미친다는 것을 보여주었다 [9,13,16].
- 최근 모델링 硏究에서는 C. arabica 蛋白質이 클로로겐산과 相互作用하기 더 쉽고 폴리페놀 酸化酵素 活性이 이 過程에서 중요한 役割을 한다는 것이 確認되었다 [17].
- 커피 品質을 包含한 食品 品質 決定에 있어 마이야르 反應(the Maillard reaction)의 重要性은 잘 알려져 있지만,
高品質 커피를 選拔/育種하기 위한 遺傳學 硏究에서는 蛋白質이 看過되어 왔다 [1,18]. - 카페인(Caffeine), 트리고넬린(trigonelline), 수크로오스(sucrose), 클로로겐산(chlorogenic acids)은 硏究에서 가장 자주 보고되는 化合物인데, 이는 定量化가 容易하고 蛋白質의 化學的 複雜性을 갖지 않기 때문일 것으로 推定된다.
- 이러한 特性을 이해하는 간단한 方法은
⇒ 커피 種子 蛋白質의 2次元 겔 電機泳動(two-dimensional gel electrophoresis)을 통해
⇒ 複雜性을 觀察하는 것이다 [8,9,19]. - Thaler & Gaigl [20]의 데이터를 기반으로 Clifford [6]는
⇒ 커피콩의 水溶性 蛋白質(water-soluble proteins)(알부민 分劃)의 아미노산 프로필(amino acid profile)이
⇒ 總 蛋白質 프로필(profile of the total proteins)과 類似하다는 사실에 주목했다. - 이러한 類似性은
⇒ 커피콩에서 가장 풍부한 蛋白質이
⇒ 11S-글로불린 種子 貯藏 蛋白質 (11S-globulin seed storage protein)이기 때문에 豫想할 수 있다 [8,21,22].

- 아미노산 타입 뿐만 아니라 炭水化物도 Maillard 反應에서 발달되는 플레이버와 아로마에 役割을 한다는 점도 고려해야 한다 [23].
- 蔗糖(Sucrose)은
커피콩의 主要 可溶性 糖이지만, 열매의 成熟 段階에 따라 果糖(fructose)과 葡萄糖(glucose)이 상당량 존재할 수 있다 [3]. - 라피노스(raffinose), 스타키오스(stachyose), 리보스(ribose), 만노스(mannose), 갈락토스(galactose)와 같은 低分子量 炭水化物도 커피콩에서 보고되었다 [15,24].
- 蔗糖 含量은 커피콩마다 크게 다를 수 있으며, C. arabica 콩의 경우 4.6~8.6 사이의 값이 보고되었다 [15].
- 커피콩은 또한 건조 중량의 40~50%를 차지하는 多糖類(polysaccharides)가 풍부하다 [25].
- 평균적으로 多糖類는
┌ 만난(mannans) 50%,
├ 아라비노갈락탄(arabinogalactan) 30%,
├ 셀룰로오스(cellulose) 15%,
└ 펙틴(pectin) 5%로 구성된다 [24]. - 低分子量 糖은 로스팅 過程에서 점진적으로 破壞된다.
- 그러나 多糖類로부터 單糖類(monosaccharides)의 形成은 유의미한 速度로 일어나지 않는 것으로 알려져 있지만, 더 복잡한 構造를 形成하는 것으로 알려져 있다 [25].
- 低分子量 糖과 마찬가지로, 온전한 多糖類와 이러한 複合 炭水化物은 遊離 아미노산, 펩티드 및 蛋白質과 反應할 수 있다. 이러한 複雜性으로 인해 각 炭水化物과 아미노산이 커피의 플레이버와 아로마 發達에 중요한 役割을 한다는 것은 알려져 있지만, 각각의 役割을 정확하게 규명하는 것은 불가능하다.
- 커피의 플레이버와 아로마 發達에 蛋白質이 관여한다는 보고가 여러 건 발표되었고,
Clifford [6,15]와 De Castro and Marraccini [26]가 이를 리뷰했다. - 그러나
11S reserve protein [8,21,22]과
다양한 커피 종에서의 貯藏 蛋白質 [27],
펩티드 [28], 그리고
프로테아제(proteases)[28]의 生化學 및 分子的 調節에 대한 몇몇 논문을 제외하고는
커피 蛋白質과 커피 品質과의 關係에 대한 硏究는 거의 수행되지 않았다. - 특정 酵素에 대한 몇몇 硏究가 발표되었지만, 品質 發達에 필수적인 것은 아니다 [16,29,30].
- 2003년에 두 가지 예외가 발표되었다.
- Montavon et al. [6]은
熟果(maturation)에 따른 蛋白質 프로파일의 變化를 보고하고 이를 컵 品質과 연관시켰으며, - Montavon et al.[15]은
로스팅 중 커피 蛋白質 프로파일의 變化에 대해 보고했다. - 우리가 아는 한, Homma [31]는
커피 아미노산과 蛋白質에 대한 章을 할애한 마지막 리뷰를 썼지만 두 페이지도 채 되지 않는다. - Flament [32]의 책에는 아로마 形成에 있어 蛋白質의 重要性이 言及되었지만 蛋白質에 대한 章이 없다.
- 최근 Hwang et al. [33]은
⇒ 커피 로스팅 중 揮發性 化合物 形成에 대한 蛋白質의 影響을 硏究했다.
⇒ 미네랄 오일 욕조를 사용하는 간단한 로스팅 裝置를 설치하여 著者는
⇒ 蛋白質이 로스팅 중 蔗糖 分解를 增加시키고
⇒ 더 많은 量의 蛋白質이 더 밝은 색상의 로스팅 커피를 生成한다는 結論을 내릴 수 있었다. - 本 章에서는 Clifford [6,15]의 리뷰 이후 발표된 最近 文獻에 특히 주목한다.
대부분의 보고는 어떤 식으로든 11S 貯藏 蛋白質(11S reserve protein)과 關聯이 있다.
또한, 프로테아제(proteases)의 役割도 論議한다.
18.2 The 11S Seed Storage Protein of Coffee
- 11S seed storage protein (種子 貯藏 蛋白質)이 C. arabica 콩으로부터 클로닝되었으며 잘 特性 分析되었다 [8,21,22].
- 선행 보고들에 따르면 SDS-PAGE 電機泳動에서 2개의 주요 밴드가 우세했으며 [12,13,34,35],
Luthe [36]는 이것이
legumin-like protein (레구민-類似 蛋白質)의 α-서브유닛과 β-서브유닛이라고 提示했다. - Acuña et al. [21]은
⇒ C. arabica 品種인 Colombia의 種子로부터 명백한 分子量이 55 kDa인 11S globulin을 分離해냈으며,
⇒ 變性 條件(SDS-PA GE)에서 22 kDa (β-arm)와 33 kDa (α-arm)의 두 폴리펩티드로 절단했다.
⇒ 分子的 特性 分析 結果 커피 레구민(coffee legumin)에는
추정 레구민 박스(legumin box)가 있는 것으로 나타났다. - Rogers et al.[8]은
⇒ C. arabica 品種 Caturra에서 抽出한 11S 커피 蛋白質을 더 잘 特性分析하기 위해
⇒ 2D-electrophoresis (電機泳動)과
아미노산 마이크로시퀀싱(amino acid microsequencing)을 사용했다.
⇒ 그들은 α- 및 β-서브유닛들로 식별된 여러 풍부한 폴리펩티드들을 시퀀싱하여
그 蛋白質이 다른 동형(isoforms)을 가지고 있음을 提示했다.
⇒ 이 폴리펩티드들을 Acuña et al. [21]에서 설명한 것과 비교한 결과,
序列 간에 23개의 아미노산이 달랐기 때문에 동형의 存在를 뒷받침했다.
⇒ 다른 植物의 11S 貯藏 蛋白質과의 類似性으로 인해, Rogers et al. [8]은
이 蛋白質이 커피에서 貯藏 技能을 가지고 있다고 提示했다.
⇒ 이 發見은 C. arabica의 씨앗에서 發芽하는 동안
α- 및 β-서브유닛의 레벨이 減少한다는 것을 보여준
Shimizu & Mazzafera [37]에 의해 추가로 確認되었다. - Rogers et al. [8,15]에 따르면,
⇒ 11S 蛋白質은 커피 胚乳(endosperm)의 總 可溶性 蛋白質의 약 45%를 차지하지만,
⇒ 胚芽(embryos)에서 發見된 蛋白質의 2D-電機泳動에서도 2개의 서브유닛들이 檢出되었다.
⇒ 胚乳에서 우세한 蛋白質이기 때문에, 클로닝된 11S 코딩 序列의 추론된 아미노산 組成은
黃-含有 아미노산(methionine 및 cysteine)이 커피콩의 총 蛋白質에 소량 존재한다는 以前 結果를
確認시켜 주었다.
⇒ Glutamate/glutamine (글루탐산/글루타민)과
aspartate/asparagine (아스파르트산/아스파라긴)이
➡ 커피 11S 蛋白質과 커피콩의 總 可溶性 蛋白質 分劃의 주요 아미노산이다. - 특정 프로모터(promoter)가 커피콩에서 11S 遺傳子의 發現을 制御한다 [22].
- Rogers et al. [8]은
시스테인과 메티오닌이 csp1에 의해 코딩되는 20개 아미노산 중 각각 1.1과 0.2에 불과하다는 것을 發見했다. - Thaler & Gaigl [20]은
메티오닌이 브라질 아라비카 커피의 總 蛋白質과 알부민 分劃에서
가장 풍부하지 않은(2.38%) 아미노산이라는 것을 發見했다. - 시스테인은 分析되지 않았다.
2개의 시스테인 殘基가 11S 커피 蛋白質의 α- 및 β-서브유닛 사이의 二黃化物 브릿지(disulfide bridge) 形成에
關與하는 것으로 提示되었다 [8]. - 黃-含有 아미노산은
커피 로스팅 중 飮料의 맛과 香 發達에 중요하기 때문에,
11S 蛋白質의 遺傳的 變異性을 調査하여
시스테인과 메티오닌 含量이 더 높은 蛋白質을 分離하는 것이 有用할 것이다 [8].
18.3 A Family of 11S Protein of Coffea
- C. arabica와
C. canephora 品種인 Robusta 콩에서 分離한 蛋白質의 2次元 電機泳動 프로파일은 서로 다르다 [8]. - 다른 펩티드들이 前驅體로 確認되었을 뿐만 아니라, α- 및 β-arms의 해당 그룹도 프로파일에 나타났다.
- 이들은 質量이 약간 더 높고 酸性 pIs가 더 높았다.
- 흥미롭게도, Robusta 品種의 모든 콩이 이러한 蛋白質 特性을 보인 것은 아니다.
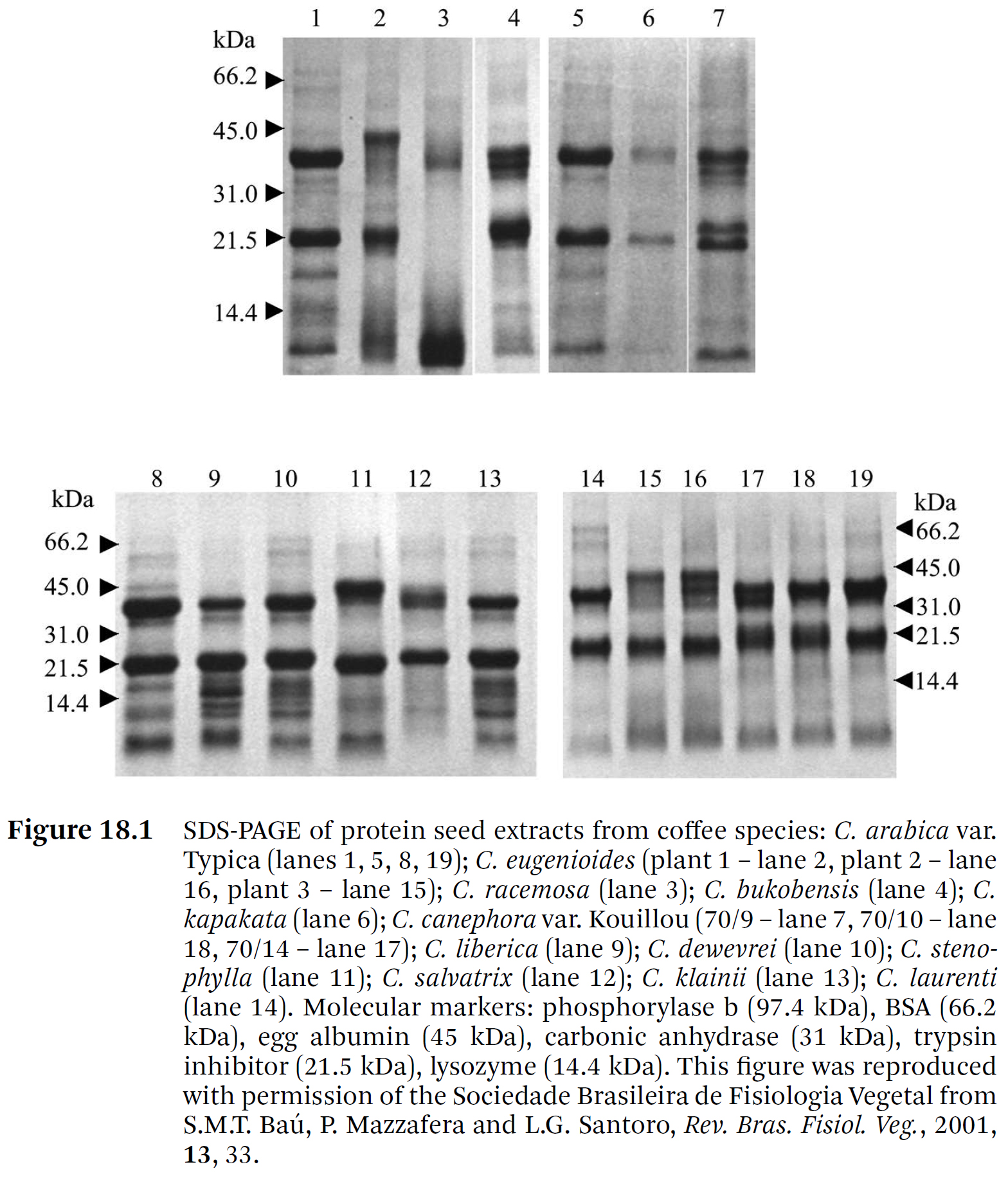
- Baú et al.[27]은
⇒ SDS 單次元 電機泳動을 사용하여 여러 C. arabica 品種들과 다른 Coffea 種의 蛋白質을 調査했다.
⇒ C. Arabica의 Typica, Maragogipe, Catuaí, Mundo Novo, Cera 및 Caturra 品種에서
각각 나무 세 그루 씩을 硏究한 결과, 매우 類似한 蛋白質 프로파일을 보였으며, 37.6 ± 3.3 및 20.9 ± 1.4 kDa의
두 밴드가 뚜렷했다.
⇒ Catuaí와 Caturra는 矮性 品種(dwarf varieties)이고,
Maragogipe는 큰 콩을 가지고 있으며,
Cera는 노란 콩을 가지고 있고,
Mundo Novo는 Typica에서 유래한 Bourbon 品種의 돌연변이이다 [38–42]. - Baú et al.[27]은
⇒ C. eugenioides와 C. canephora variety Kouillou (또는 Conillon)의 세 나무들과,
C. racemosa, C. bukobensis, C. kapakata, C. liberica, C. dewevrei, C. stenophylla,
C. salvatrix, C. klainii, C. laurentii의 각 나무 한 그루를 分析했다 (Figure 18.1).
⇒ 蛋白質 含量의 變化 외에도,
C. racemosa의 4.7%에서 C. liberica의 9.1%로
蛋白質 프로필, 즉 두 서브유닛의 質量과 서브유닛의 數 모두에서 큰 변차가 觀察되었다.
⇒ 예를 들어, 세 그루의 C. eugenioides 나무 중 plant 2 (레인 16)에는 추가 α-서브유닛이 있었다.
⇒ C. canephora var. Kouillou 나무 70/9 (7번 레인)는
분명히 추가적인 α- 및 β-서브유닛을 가지고 있었다.
⇒ C. Racemosa (2번 레인) 프로파일은 가장 달랐는데, 약 12 kDa의 강한 밴드를 보였다.
⇒ 또한, α-arm 위치에서 밴드가 檢出되었지만,
β-arm의 겉보기 質量에서는 밴드가 觀察되지 않았다. - Bandil et al.[43]은
⇒ 두 가지 다른 發達 段階에서 C. arabica 콩의 2D 電機泳動 프로파일을 生成했다 :
⇒ 1단계는 細胞分裂과 胚乳 周圍의 伸張이고,
⇒ 2단계는 胚乳의 初期成長이 外胚乳(perisperm)를 점진적으로 代替하는 段階이다.
⇒ 2段階에서는 6개의 蛋白質만 差等的으로 發現되었는데,
그 중 4개는 에너지 代謝와 關聯이 있었고,
하나는 타우마틴(thaumatin),
다른 하나는 11S로 確認되었다.
18.4 2S Protein of Coffea
- Lin et al. [44]은
C. canephora 열매에서
⇒ 2S-type 蛋白質을 코딩하는 遺傳子를 優性 轉寫體(dominant transcript)로 確認했다.
⇒ 두 번째로 많이 發現되는 遺傳子는 11S reserve protein과 關聯이 있었다.
⇒ 이 硏究에서는 잎, 果皮, 씨앗과 果皮가 섞인 어린 체리, middle-stage 씨앗 및 late-stage 씨앗의
라이브러리를 기반으로 EST 데이터베이스를 구축했다.
⇒ 흥미롭게도 2S 遺傳子는 열매 成熟 중에 一時的 發現을 보였다 [21] ;
ESTs는 early stage에서,
1037개는 middle stage에서,
139개는 late stage에서 發見되었으며,
⇒ 이는 전체 데이터베이스의 98.2% (1219개 ESTs 중 1197개)를 차지했다.
⇒ 그러나 11S 遺傳子는 early stage (28) 및 middle stage (244)에 비해
late stage(412)에서 가장 높은 轉寫體 數를 보였다.
⇒ 2S 遺傳子는 토마토, 참깨, 해바라기, 브라질 너트의 2S 蛋白質과
가장 높은 類似性을 보였다 (類似性이 높은 것부터 순서대로). - Mondego et al. [45]는
⇒ Lin et al. [44]이 수행한 C. canephora 硏究에서 얻은 8개의 EST 라이브러리와
브라질 커피 게놈 프로젝트[46]에서 얻은 43개의 C. arabica 라이브러리를 結合했다.
⇒ 브라질 프로젝트의 소스는
주로 두 가지 C. arabica 品種 (cv. Rubi의 發芽 種子를 除外한 Catuai와 Mundo Novo)과
한 가지 C. canephora 遺傳子型(Conillon)이었다. - Lin et al. [44]의 硏究에서
⇒ 소스는 다섯 가지 다른 C. canephora 遺傳子型이었다.
⇒ C. Canephora와 마찬가지로
참깨의 2S 蛋白質과 相同하는 2S 遺傳子가 C. arabica에서 높게 發現되었다.
⇒ 이 遺傳子는 다섯 번째로 많이 發現되는 遺傳子였으며,
4개의 라이브러리에서 發見되었고 1271개의 EST로 表現되었다.
⇒ 11S 遺傳子는 C. arabica에서 가장 많이 發現되는 20개 遺傳子에 包含되지 않았다.
⇒ 안타깝게도 브라질 커피 게놈 프로젝트에는 열매의 發達 段階를 구분하는
열매 전용 라이브러리가 없었다.
⇒ FR1과 FR2 라이브러리는
Mundo Novo에서 꽃봉오리 + 핀헤드 열매 + 각기 다른 단계의 열매로 기술되었다
(FR1과 FR2 라이브러리).
⇒ 2S 蛋白質과 關聯된 대부분의 ESTs는 이 라이브러리에서 發見되었다. - C. racemosa는 브라질 EST 프로젝트에서도 硏究되었다 [46].
⇒ 열매들에서 抽出한 두 라이브러리는 “FR4–fruits”와 “FV2 – stages 1, 2, 3"로 기술되었다.
⇒ C. racemosa의 EST에 대한 追加 情報는 없지만,
브라질 EST 프로젝트(http://bioinfo03.ibi.unicamp.br/cafe/)에서
공개적으로 이용 가능한 Electronic Northern을 檢索한 결과,
⇒ 2S 遺傳子가 가장 많이 發現되는 20개 遺傳子 중 하나였으며
참깨의 2S 蛋白質과 相同性을 보였다.
⇒ 11S 遺傳子는 C. racemosa의 EST에서 發見된 遺傳子에 包含되지 않았다.
⇒ 다른 더 많이 發現되는 遺傳子는 주로
┌ 防禦(chitinases, chaperones, lipid transfer proteins),
├ 光合成(Rubisco small unit, chlorophyll a/b binding protein),
├ 酸化還元 反應 調節(catechol oxidase),
├ 燐酸 代謝(acid phosphatase가 가장 많이 發現되는 遺傳子),
└ 물 輸送(aquaporins)과 같은 過程과 關聯이 있었다. - 참깨 2S reserve protein의 分子量은 13~15 kDa이며 [47],
⇒ 이는 C. racemosa의 電機泳動 프로파일 (Figure 18.1 - Baú et al. [27]) 및
브라질 EST 프로젝트(Mazzafera, 未公開 資料)에서 發見된 뉴클레오티드 시퀀스의
번역된 오픈 리딩 프레임의 주요 蛋白質 質量과 類似하다.
⇒ 이 EST 데이터베이스의 序列 分析 결과,
C. racemosa에는 두 가지 同型蛋白質(Mazzafera, 未公開 資料)가 存在하는 것으로 나타났으며,
이는 Coelho et al.[48]의 결과와 一致한다.
⇒ 이 著者들은
C. arabica와 C. racemosa의 成熟한 種子에서 두 段階의 크로마토그래피를 통해
legumin-like proteins를 정제하고,
각 種에서 각각 CaL-1과 CaL-2, CrL-1과 CrL-2라는 두 개의 分劃을 分離했다.
⇒ 變性 電機泳動(Denaturing electrophoresis) 결과,
CaL-1은 33 kDa와 24 kDa의 2개의 뚜렷한 밴드로 構成되어 있음을 確認했다.
CaL-2, CrL-1, rL기-2는 apparent molecular mass가 14 kDa인 單一 蛋白質 밴드를 보였다.
그러나 CaL-2, CrL-1, CrL-2의 아미노산 組成은 상당히 달랐다. - Coelho et al. [48]은
⇒ 인공 사료를 사용한 用量-反應 實驗에서
⇒ Callosobruchus maculatus의 幼蟲 發達이 統計的으로 有意味하게
減少하고 사망률이 增加하는 것을 觀察했기 때문에,
⇒ C. racemosa의 2S legumin-like protein이
이 種의 coffee berry borer Hypothenemus hampei [49]에 대한 抵抗性에 關與할 수 있다고 提示했다.
⇒ 이러한 측면에서, C. arabica가 이 昆蟲에 민감한 이유는
C. racemosa의 2S 蛋白質과 比較했을 때
2S 蛋白質의 아미노산 序列이 다르고 種子에서 發見되는 量이 적기 때문일 수 있다. - 또한, Lin et al.의 EST 硏究 [44]는
⇒ H. hampei에 취약성이 있는 C. canephora 콩의 成熟 過程에서
2S 轉寫體가 減少하고 11S 轉寫體가 增加함을 보여주었다.
18.5 Peptides and Proteases
- 여러 타입의 커피에서 遊離 아미노산의 組成이 硏究되었으며, 아로마 前驅體로서의 役割은 잘 알려져 있다.
- 펩티드는 이와 같은 關心을 받지 못했다.
- Ludwig et al. [28]은
⇒ 여러 C. arabica 샘플에서 평균적으로 펩티드가 乾燥 重量의 약 0.59%,
C. canephora 샘플에서 0.43%임을 發見했다.
⇒ 아미노산과 蛋白質이 提擧된 샘플을 2D-PAGE로 分離하여
分子量이 4~10 kDa인 30개의 펩티드를 識別했다.
⇒ C. arabica와 C. canephora 빈들은
2D 電機泳動 프로파일이 달랐으며,
total protein 및 11S reserve proteins와 마찬가지로
메티오닌(methionine)이 펩티드들 중 가장 풍부하지 않은 아미노산이었다.
⇒ ┌ Glutamic acid/glutamine,
├ aspartic acid/asparagine 그리고
└ glycine은
두 종류의 커피 모두에서 가장 풍부한 아미노산이었다.
⇒ 펩티드는 5명의 센서리 느낌을 이용한 모델 로스팅 실험에서 덱스트로스(dextrose)와 함께 녹였고,
커피의 아로마 形成에 있어서의 役割을 確認했다. - Ludwig et al. [28]은
⇒ 또한 겔 電機泳動 후 活性 染色을 사용하여
커피 蛋白質 抽出物의 蛋白質 分解 活性을 檢定했고 모든 커피 샘플에서 최소 5~7개의 活性 밴드를 發見했다.
⇒ 이들의 活性은 세린(serine) 및 펩신 프로테아제(pepsin protease) 抑制劑에 의해 減少되지 않았지만,
요오드아세트아미드(iodoacetamide)는 명확한 減少를 招來했다.
⇒ 매우 중요한 觀察은 蛋白質 分解 活性이 貯藏된 커피 콩에서 수개월 동안 安定的이라는 事實이었다.
⇒ 그러나 1년 동안 貯藏된 원두의 아미노산 프로필은
글루타민(glutamine)의 減少를 제외하고는
總 蛋白質 아미노산 含量(total content of protein amino acids)이나
⇒ 대부분의 個別 아미노산 濃度를 크게 變化시키지 않았다는 점이 흥미롭다 [50]. - 따라서 커피의 11S 蛋白質이 發芽 중에 分解되기 때문에
蛋白質分解 活性(proteolytic activity)이 觸發되어야 하는 것으로 보인다 [37]. - Montavon et al. [9]은
⇒ 분말 콩의 無酸素 培養(anoxic incubation)이 아닌
有酸素 培養(aerobic incubation)이
11S reserve protein의 斷片化(fragmentation) 또는 消化(digestion)와
遊離 아미노산의 放出을 促進한다는 것을 觀察했다.
⇒ 有酸素 條件에서 11S reserve protein의 α-subunit은
抽出物에서 즉시 사라졌지만 β-arm은 부분적으로 保存되었다.
⇒ 이러한 結果는
蛋白質分解 消化(proteolytic digestion)보다는 酸化 過程(oxidative process)을 시사하지만,
⇒ 著者들은
酸化가 蛋白質 構造를 變化시키고
活性 蛋白質 分解 酵素(active proteases)에 대한 敏感度를 增加시킬 수 있다고 주장했다. - Lepeley et al. [51]는
⇒ C. canephora 콩에서 두 가지 시스테인 프로테아제(cysteine proteases)의
遺傳子 序列(gene sequences)을 分離하여
열매 熟果(fruit maturation)에 따라 發現(expression)이 增加함을 보였다.
⇒ 그러나 CcCP1의 發現 水準은 CcCP4의 發現 水準보다 600% 더 높았다. - Abreu et al. [52]은
⇒ 브라질 EST 프로젝트 데이터베이스에서 프로테아제 시퀀스들을 檢索했다.
⇒ 그들은 C. arabica 組織으로 만든 모든 라이브러리에서
41개의 遺傳子(콘티그)를 찾았고,
蛋白質 도메인을 基盤으로 34개가 시스테인 프로테아제(cysteine proteases),
6개가 아스파르트산 프로테아제(aspartic proteases), 그리고
1개만 세린 프로테아제(serine protease)로 確認되었다.
⇒ 14개의 遺傳子는 열매 라이브러리에만 存在했으며,
3개의 시스테인 프로테아제(CaCP1, CaCP4, CaCP23)와
1개의 아스파르트산 프로테아제(CaAP2)를 追加 硏究를 위해 選擇했다.
⇒ CaCP4, CaCP23 및 CaAP2의 發現 프로파일은
成熟 열매보다 未成熟 열매에서 더 많이 發現되는 반면, CaCP1의 경우 반대 경향이 觀察되었다 [52].
⇒ 번역된 序列로 구축된 系統樹(Phylogenetic trees)는
CaCP1과 CaCP4가 각각 CcCP1 및 CcCP4와 매우 類似함을 보여주었다.
⇒ CaCP1, CaCP4 및 CaCP23은 서로보다 다른 種의 蛋白質 分解 酵素와 더 밀접한 關聯이 있었다.
⇒ CaAP2는 C. canephora에서 分離된 CcAP2와 매우 類似했으며 [53],
둘 다 CcAP1과 완전히 달랐다. - 브라질 EST 프로젝트 데이터베이스에서
시스테인 프로테아제 序列이 우세하고 未成熟 열매에서 가장 높게 發現된다는 사실은
activity assays와 일치한다 [52].
⇒ 겔 活性 分析(gel activity assays) 결과,
未熟豆에서 얻은 蛋白質 抽出物에서 가장 높은 活性이 發見되었으며,
요오드아세트아미드(iodoacetamide) 처리 후 이 活性이 강하게 抑制되는 것으로 나타났는데,
이는 Ludwig et al.[28]의 결과를 뒷받침한다. - 흥미롭게도 Abreu et al. [52]도
⇒ 세린 프로테아제 活性(serine protease activity)을 검출했는데,
⇒ 이는 페닐메틸설포닐 플루오라이드 (phenylmethylsulfonyl fluoride, PMSF)에 의해 抑制되었지만,
커피 EST 데이터베이스에서 프로테아제를 검색하는 동안
세린 프로테아제 遺傳子를 確認하지 못했다.
⇒ 이러한 불일치를 설명하기 위해
이러한 硏究에 사용된 品種의 特性, 조사된 發達 단계 및 열매 收穫 場所가 논거로 사용되었다. - Abreu et al. [52]은
⇒ 브라질의 두 곳에서 샘플을 수집하고,
프로테아제 活性 및 關聯 遺傳子 發現 외에도
蛋白質 및 遊離 아미노산 含量을 分析하여 아미노산 프로파일을 生成했다.
⇒ 샘플은 상파울루 주의 아다만티나(Adamantina)와 모코카(Mococa)에서 蒐集되었는데,
두 地域은 氣溫 差異가 뚜렷하고 독특한 品質의 커피를 生産한다.
⇒ 모코카는 서늘한 地域으로 커피 品質이 더 좋다.
⇒ 전반적인 결과는 높은 기온이 낮은 品質과 높은 프로테아제 活性과 關聯이 있음을 나타냈다.
⇒ 이 결과는 같은 지역에서 灌漑의 影響을 조사한 선행 보고서[54]에서도 뒷받침된다. - 4가지 灌漑 處理를 거친 C. arabica 나무의 蔗糖, 還元糖, 澱粉, 總 페놀性 物質, 클로로겐산(5-카페오일퀸산), 카페인, 脂質, 蛋白質, 遊離 아미노산 및 窒素 含量과 프로테아제 및 폴리페놀 酸化酵素의 活性에 대한 主成分 分析(PCA) 결과, 灌漑보다는 溫度가 品質에 影響을 미치는 主要 要因인 것으로 나타났다.
18.6 Does Coffee Have Bioactive Proteins and Peptides?
- 生理活性 蛋白質과 펩티드는
⇒ 窒素源으로서의 役割 外에도
⇒ 生理學的 重要性에 대해 많은 食品 供給源에서 硏究되어 왔다. - 우유, 옥수수, 생선, 밀, 쌀, 대두 등 여러 食品 供給源에서 發見될 수 있으며 [55,56],
肯定的 및 否定的 生理的 效果의 範圍가 매우 넓다 [57]. - 蛋白質과 펩티드의 生理活性的 役割과 營養學的 影響에 대한 情報는 지난 몇 년 동안 상당히 增加했다 [58].
- 蛋白質의 크기 때문에,
손상되지 않은 蛋白質은 人間의 胃腸管에 吸收되지 않아 生理的 作用이 制限된다. - 그러나 다른 식품 공급원에도 존재하는 펩티드와 蛋白質 消化로 生成되는 펩티드는
더 작고 吸收될 可能性이 높아 다양한 生利活性을 나타낼 수 있다 [58]. - 안타깝게도 機能性과 構造的 特性 간의 關係에 대한 知識은 아직 不足하다.
- 커피의 蛋白質과 펩티드의 生理活性에 관한 情報는 없다.
- Farah [59]는
카페인, 트리고넬린, 클로로겐산, 테르펜, 纖維質, 지질, 미네랄, 탄수화물 및 其他 化合物을 包含하여 커피의 여러 生理活性 成分을 나열했지만, 蛋白質과 펩타이드는 包含되지 않았다. - 커피는 抽出 前에 로스팅되므로 蛋白質 프로필이 크게 변경된다 [9,10].
⇒ 커피를 로스팅하는 데 사용되는 온도에서는
蛋白質이 전체 또는 부분적으로 破壞될 뿐만 아니라
11S 커피 貯藏 蛋白質에서 觀察된 것처럼 펩티드가 放出된다 [10]. - 몇몇 “omic" 스터디들을 包含하는 "푸드오믹스"의 글로벌 觀点을 기반으로,
蛋白質 "digestomics (消化體學)"은 식품에서 蛋白質과 펩티드의 機能을 硏究하는
새로운 플랫폼으로 提案되었다 [60]. - 커피 硏究는 이 이니셔티브를 통해 이익을 얻어야 한다.
- 최근 출간된 책에서는 커피 成分의 利點과 危險을 포괄적으로 다루었지만
蛋白質과 펩티드에 대한 章은 없었다 [61]. - 따라서 커피 蛋白質 硏究에는 공백이 있는 것이 분명하며,
本 章에서 인용된 프로테오믹스(proteomic) 硏究를 통해
生成된 知識을 擴張하고 이러한 노력을 健康 硏究와 결합해야 한다.
18.7 Conclusion
- 많은 보고서에서
로스팅된 커피 원두의 아로마와 플레이버 形成에
蛋白質, 펩티드, 아미노산의 중요성을 강조했으며, 이는 부인할 수 없는 사실이다. - 그러나 이러한 情報는 不足하며 대부분 成熟한 콩들을 사용하여 얻어졌다.
- 이용 가능한 誘電體 情報[44–46]와
잘 계획된 프로테옴 實驗(proteome experiment)의 데이터를 연관시키는 것이
커피 콩의 蛋白質을 識別하는 戰略으로 사용되어야 한다. - 이상적으로 이러한 實驗은
컵 品質에 현저한 차이를 보이는 品種의 C. arabica 콩을 사용하거나,
Abreu et al. [52]과 Silva et al. [54]의 硏究에서 提示된 것처럼
氣候條件에 의해 品質이 影響을 받는 최소 두 곳에서 재배된 동일 品種의 C. arabica 콩들을
사용하여 수행해야 한다. - 안타깝게도 Bandil et al. [43]이 수행한 프로테옴 硏究는 두 가지 發達 段階만 分析했다.
- 따라서 제안된 硏究에서는
成熟에 가까운 여러 단계의 열매에서 抽出한 콩들과
最適 및 最適이 아닌 條件에서의 貯藏이 品質 低下를 誘發하는 影響을 考慮해야 한다. - 또한, 프로테아제 活性度도 追跡해야 한다.
- 이러한 實驗이
커피 콩 總 蛋白質의 45%를 차지하는 11S 蛋白質과 같은 蛋白質이
커피 아로마와 플레이버 發達에 어떻게 關與하지에 대한 모든 의문에 답할 수는 없겠지만,
이용 가능한 情報를 戀結하고
커피 品質 硏究의 새로운 方向을 提示할 수 있는 탄탄한 基盤을 提供할 것이다. - 2S 遺傳子는 커피 콩 熟果 過程에서 고도로 發現되므로 2S 蛋白質 또한 고려해야 한다.
- 炭水化物이 이러한 狀況에서 중요한 役割을 한다는 점도 認知하고 있으며,
이에 대한 竝行 硏究도 遂行해야 할 것이다. - 또한, 커피 蛋白質과 펩티드의 生理的 效果를 규명하기 위한 노력도 필요하다.
- 적당한 양의 커피 攝取는 人體 健康에 도움이 된다고 알려져 있으며,
다른 많은 食品과 마찬가지로 이러한 效果의 일부는 이러한 化合物과 關聯이 있을 수 있다.
References






'Coffee Chemistry' 카테고리의 다른 글
| 커피의 窒素化合物 (12) | 2025.07.28 |
|---|---|
| 커피의 脂質 (21) | 2025.07.25 |
| 커피 멜라노이딘의 분석, 구조, 형성경로, 생리활성 및 건강영향 (16) | 2025.07.23 |
| 커피 멜라노이딘 : 구조, 형성, 건강 효능 (16) | 2025.07.20 |
| 커피의 화학 성분 (17) | 2025.07.17 |




댓글