
- 우리는 野生 아라비카 커피(Coffea arabica)의 生物學的 起源과 進化에 대한 정확한 세부 사항을 결코 알 수 없지만, 그 식물과 근연종들에 관한 연구를 기반으로 몇 가지 그럴듯한 추론을 할 수 있다.
- 광범위한 과학 연구를 통해 우리는 아라비카 커피가 두 가지 아프리카 커피 種인
⇒ Robusta coffee (Coffea canephora)와
⇒ Nandi coffee (C. eugenioides)의 交配 結合(hybrid union)에서 나왔거나, 또는
아마도 매우 밀접하게 관련된 다른 두 種(현존 또는 멸종)의 交配에서 생겼다는 것을 알고 있다. - 로부스타 커피는 많은 열대 아프리카 지방에 널리 分布되어 있다.
난디 커피는 주로 아프리카 리프트 밸리 지역의 중부와 북부의 고지대와 리프트 밸리 동쪽의 일부 지역에서 發見된다. - 이 두 종의 交雜(hybridization)이 어디에서 일어났는지는 알려져 있지 않다.
하지만 에티오피아와 가까운 지역에서 발생했거나 에티오피아 자체에서 發生했을 可能性이 가장 높다. - 오늘날 야생 로부스타와 난디 커피는 남수단, 콩고민주공화국, 케냐, 탄자니아, 우간다에서 함께 자라고 있거나 서로 가까이에서 發見된다 (Davis et al. 2006).
- 그러나 최근의 地質學的 時間 동안 기후가 변했기 때문에 이 두 種의 分布는 오늘날 수천년 전과 다를 가능성이 높다.
- 실제로 로부스타와 난디 커피는 地質學的 過去의 어느 時點에 에티오피아에 存在할 수 있었다.
- 로부스타와 난디 커피의 異種交配(hybridization)는
異質倍數性(allopolyploidy)으로 알려진 현상인
染色體數의 重複(duplication of the number of chromosomes)로 이어졌다. - 아라비카 커피에서는 染色體가 두 배가 되었기 때문에,
다른 모든 커피 種에서와 같이 22개의 염색체를 갖는 대신 44개의 染色體가 있다. - 아라비카는 4세트의 染色體를 가지고 있기 때문에 異質四倍體(allotetraploid)로 분류된다.
- Allopolyploidy와 관련된 결과가 있는데, 어떤 것은 좋고 어떤 것은 나쁘다.
- Allopolyploidy는, 染色體數의 증가 없이 이종 교배(hybridisation) 만으로도 하는 것처럼,
식물을 더욱 활기차게 만들고 더 넓은 범위의 環境條件에 더 잘 適應할 수 있게 한다. - 보편적이지는 않지만 allopolyploidy의 또 다른 결과는 自家受精 能力(ability to self-fertilise)이다.
자가수정 식물에서는 꽃의 수정에 他家受粉(cross pollination)(즉, 다른 식물과의 교배)이 필요 없이,
생존가능한 과실과 씨앗을 생산할 수 있다. - 이것은 농작물에 유용한 속성이지만, 야생 및 경작 식물 모두에 심각한 장기적 결과를 초래한다.
- 식물이 自家受精하면
⇒ 다른(유전적으로 다른) 개체로부터 遺傳物質을 받지 않으므로,
⇒ 시간이 지남에 따라 遺傳的 多樣性(genetic diversity)이 심각하게 감소할 수 있다 (近親交配, inbreeding). - 이것은 아마도 에티오피아의 야생 아라비카 커피의 경우였을 것이다.
- 특히 초기 交配 이벤트 이후 進化生活의 초기 기간 동안 out-crossing의 기회가 매우 制限되었을 것이다.
- 더욱이, 어떤 연구들은 로부스타와 난디 커피의 交配와 異質四倍體(allotetraploid) 아라비카 커피의 형성이
단 한 번만 일어났을 수 있으며 (Tesfaye et al. 2007), 遺傳的 交換을 더욱 制限할 수 있었을 것이라고 추론한다.


- 아라비카 커피는 적어도 16世紀 중후반부터 에티오피아 밖의 과학 및 의학계에 알려졌다.
그러나 1753年이 되어서야 定式 學名이 Coffea arabica로 指定되었다.
그것은 스웨덴의 식물학자, 의사, 동물학자인 Carl Linnaeus (1707–1778)에 의해 命名되었다.
이것이 學名에 Linnaeus의 약어인 'L.'이 붙는 것을 때때로 보는 이유이다 (즉, Coffea arabica L.). - 물론 이 植物은 16세기 이전에 과학계 외부, 특히 에티오피아와 예멘에서도 잘 알려져 있었고
이미 아프리카와 아라비아 반도 외부에서 栽培되고 所費되고 있었다. - Carl Linnaeus와 그 이전의 식물학자들, 심지어 19세기 초반과 중반까지 일했던 사람들조차
커피나무가 예멘(Arabia felix; 또는 라틴어로는 Arabia felici)에서 유래했다고 믿었기 때문에
‘Arabica’라는 이름이 붙었다. - 19世紀 중후반이 되어서야 이것이 실제로 아프리카에서 온 식물이라는 사실이 과학계에 알려졌고,
훨씬 더 최근에 와서야 진정한 生物學的 故鄕이
에티오피아 하이랜드와 남수단의 숲이라는 사실이 밝혀졌다 (Davis et al. 2006). - 로부스타 커피(C. canephora)는
비교적 최근인 20世紀 初부터 과학계에 알려졌지만
우간다에서 栽培 및 使用(chewing coffee로)이 1860年代에 기록되었으며
아라비카 커피와 마찬가지로 아마도 수천년 동안 地域 住民들이 使用했을 것이다. - 코페아屬(Coffea)에 속하는 것으로 공식적으로 인정된 것들은
124種이 있으며,
아프리카, 마다가스카르 및 기타 서부 인도양 제도,
아시아 및 오스트랄리아 全域에서 發見된다 (Davis et al. 2006, 2011). - 에티오피아에서는 두 種이 자연적으로 발생한다 (즉, 야생으로 자란다).
⇒ 아라비카 커피(C. arabica) 및
⇒ C. neoleroyi. - Coffea neoleroyi는
에티오피아 남서부의 600~750m 고도에서
계절적으로 건조한 숲(Combretum-Terminalia woodland and wooded grassland (CTW))에서 발생하는
작은 낙엽 관목 또는 작은 나무(a small deciduous shrub or treelet)이다 (Friis et al. 2015).
남수단 동부와 우간다 북부에서도 발생한다 (Davis et al. 2023). - Coffea neoleroyi는 稀貴種으로 식물학자들에게도 거의 보이지 않고 거의 알려지지 않았으며
커피 생산에 사용되지 않는다. - 아라비카 커피(C. arabica)는
⇒ 에티오피아와 남수단에서 自生하지만,
남수단에서는 에티오피아 국경에 가까운 보마 고원으로 제한된다.
⇒ 케냐 북부의 Marsabit 山이 종종 야생 아라비카의 서식지로 제시되지만,
연구에 따르면 이 산에서 자라는 아라비카 나무는 경작지에서 유래했을 가능성이 있으며,
환경과 위치가 자연 발생과 일치하지 않는 것으로 나타났다 (Davis et al. 2012).
이 지역에서 아라비카 식물의 상태를 완전히 평가하기 위해서는
Mount Marsabit에서 추가 연구가 필요하다.
⇒ 다양한 문헌 출처에서 남수단 남부의 이마통 산맥(Imatong Mountains)이
아라비카 커피의 야생 개체군이 서식하는 장소라고 하지만,
이 산맥에서 유일하게 알려진 기록은 이마통에서 야생으로 자라는
로부스타 커피(C. canephora)일 가능성이 높다.
- 野生 아라비카의 가장 크고 다양한 個體群은
⇒ 에티오피아 南西部의 숲이 우거진 高地帶에서 發生한다 (Senbeta et al. 2007)(Figure 1).
⇒ 그 개체군은 Jimma-Limu, Kaffa, Bench Maji, Tepi, Illubabor 및 Wellega의 서부와
같은 커피지역에서 發見된다 (Figure 1; Figure 2).
⇒ Rift Valley의 동쪽에도 野生 個體群이 있다.
그러나 이들은 Bale 커피 지역의 숲이 우거진 지역(Figure 2)과 특히 Harrena forest로 제한된다.
⇒ Bale의 個體群은 土着은 아니지만(즉, 원래 이 지역에서 야생이 아니었음),
the Rift Valley의 동쪽에 있는 다른 지역, 아마도 Sidamo에서 이곳으로 옮겨온 것으로 추정된다.
⇒ 그러나 분자(DNA) 기술을 사용한 연구는, 아라비카 커피가 그곳의 천연식생의 일부인데,
이는 Bale Mountains(베일 산맥)에 있는 포레스트들이 남서부 에티오피아의 多濕한 (커피) 포레스트들과
매우 유사한 植物 種 構成을 가지고 있다는 사실로 뒷받침된다. - 에티오피아에서 야생 아라비카는 아틀라스 지도에서
짙은 녹색으로 표시된 excellent suitability 또는 good (niche) suitability를 가진 숲이 우거진 에리어들에서 발견된다. 예외적으로 Sidamo 커피 에리어의 西部 地域은 대부분 稠密한 混農林業(dense agroforestry)으로 덮여 있다. 그러나 Sidamo 커피 栽培地域에서 현재 栽培되고 있는 커피의 일부 또는 대부분은 원래 이 지역에 한때 존재했던 野生 숲에서 온 것일 수 있다. - 에티오피아의 野生 커피의 기록 위치를 보여주는 地圖는 Davis et al. (2012)에서도 찾아 볼 수 있다.



- 에티오피아와 남수단에서, 野生 아라비카 커피는
⇒ 해발 950~1,950m의 습한 常綠山地 숲 (moist (humid), evergreen montae forest)에서
⇒ 작은 下層部 나무(a small understory tree)로 발생하는데,
⇒ 주로 1,200m 이하 高度에서의 숲 損失 때문에,
⇒ 1,200m가 현재 實質的인 高度限界이기는 하다. - 野生 아라비카 커피는
⇒ 多濕한 常綠 아프로몬탄 숲(Moist evergreen Afromontane forest (MAF))과
⇒ 轉移 雨林(Transitional rain forest (TRF))에서 發生한다 (Figure 5) (Friis et al. 2010). - MAF 및 TRF 숲은
⇒ 다양한 종류의 常綠樹(evergreen trees)와 灌木(shrubs)으로 구성되어 있으며,
⇒ 冷-熱帶 雨林(cool-tropical rainforest)으로 가장 잘 설명되는 곳을 나타낸다. - 때때로 '커피 숲(coffee forest)'이라고도 하는 MAF 및 TRF 포레스트 타입은
⇒ Figure 1과 같이 대부분 에티오피아 하이랜드의 南東部 및 南部 地域으로 제한된다.
⇒ 이러한 숲 유형의 대부분이 農業과 定着을 위한 土地를 제공하기 위해 除去되었기 때문에,
잠재적인 커버 면적은 현재보다 훨씬 더 컸을 것이다. - MAF 및 TRF의 潛在的 森林 面積, 즉 농업 등이 아닌 경우 이러한 森林 類型이 자랄 수 있는 곳은 Figure 5에 나와 있다.
- 이러한 森林 類型의 실제 남은 面積(통칭하여 humid forest)은 Figure 1과 여타 전문 지도들에 나와 있다.
- 아라비카 커피는
⇒ 주로 MAF 및 TRF 地域에서 栽培되지만
⇒ 일부 지역, 특히 Harar 커피 지역(Arsi, West Hararge 및 East Hararge),
Amhara 지역 및 c. 2,000 m 이상의 에티오피아 남동부에 있는 하이랜드 에리어 같은 곳에서는,
高-高度 常綠樹林의 多濕한 地域에서도 栽培된다
(Dry evergreen Afromontane forest and grassland complex (DAF); Figure 5). - 野生 아라비카 커피는 에티오피아의 커피 生産에서 核心的인 役割을 한다.
⇒포레스트 및 세미-포레스트 생산 시스템에서 收穫되는 作物로서 뿐만 아니라,
⇒ 에티오피아의 많은 커피 生産地域의 植栽 및 再植栽를 위한 種子와 苗木의 源泉으로도 使用된다. - 에티오피아의 coffee forest은 또한
⇒ 야생 아라비카 커피 遺傳資源의 主要 貯藏庫로서 막대한 價値를 지니고 있으며,
⇒ 이는 疾病 抵抗性 栽培品種의 源泉으로 사용되고
⇒ 種의 컵 프로필 多樣性(예: 독특하고 우수한 맛 프로필)을 改善하는데 사용되어오고 있다. - 에티오피아의 野生 아라비카 個體群의 遺傳的 多樣性(genetic diversity)은
세계 다른 곳에서 作物 生産에 사용되는 아라비카 커피의 栽培 變種들(cultivated variants)을 훨씬 凌駕한다. - 土着 個體群들은 종종 전 세계 아라비카 生産의 中長期 持續可能性을 위한 核心 資源으로 강조된다.
- 에티오피아의 多濕한 常綠樹林(humid evergreen forests)은
生物多樣性 保存(biodiversity conservation)을 위한 세계적으로 중요한 핫 스팟으로 인식되고 있으며,
生物多樣性 保存을 위한 세계적으로 중요한 34개 優先順位 地域 중 하나인
Eastern Afromontane Biodiversity Hotspot (Mittermeier et al. 2004)의 一部이다. - 1,500m 이상 아프로몬타네 地域의 50% 이상과
2,000m 이상 아프로몬타네 지역의 약 80%가 ⇒ 에티오피아 내에서 發見된다는 점을 고려할 때,
에티오피아 하이랜드들에 있는 ‘coffee forests' 및
기타 傳統的인 커피 生産시스템은 生物多樣性 保全을 위해 국가적으로나 세계적으로도 重要하다.



- 아라비카 커피는
⇒ 작은 常綠樹(small evergreen tree)이다. (灌木(shrub)이 아님)
⇒ 높이 7m, 너비 3m 정도까지 자랄 수 있다. - 성숙기에 아라비카 나무는
⇒ 외형이 다소 삼각형(triangular in outline)이고 3차원 모양이 원추형(conical)이다.
⇒ 이는 나무 밑동(base)에 가장 긴㈜ 가지가 있고 위로 갈수록 가지가 점차 짧아지기 때문이다.
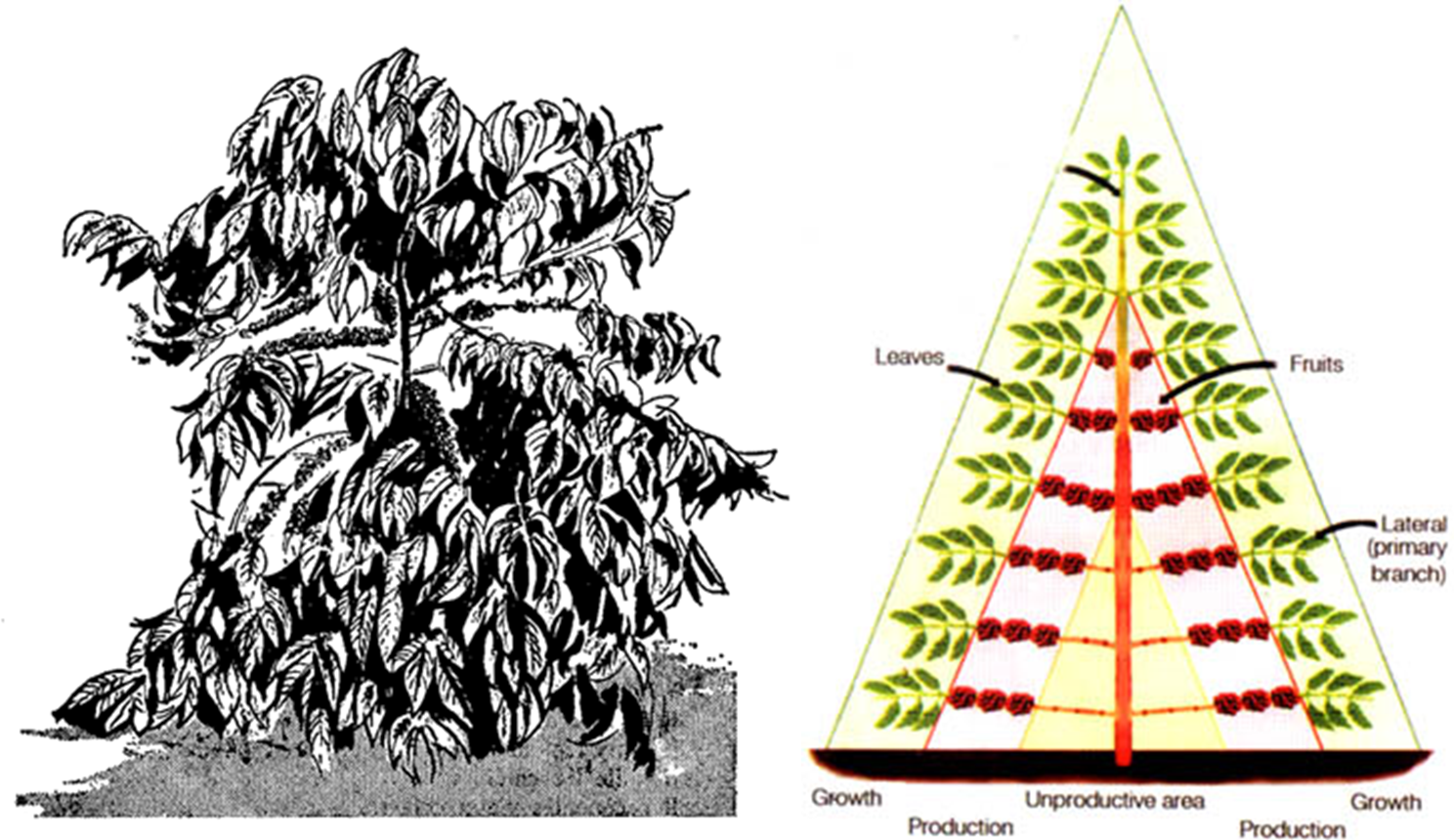
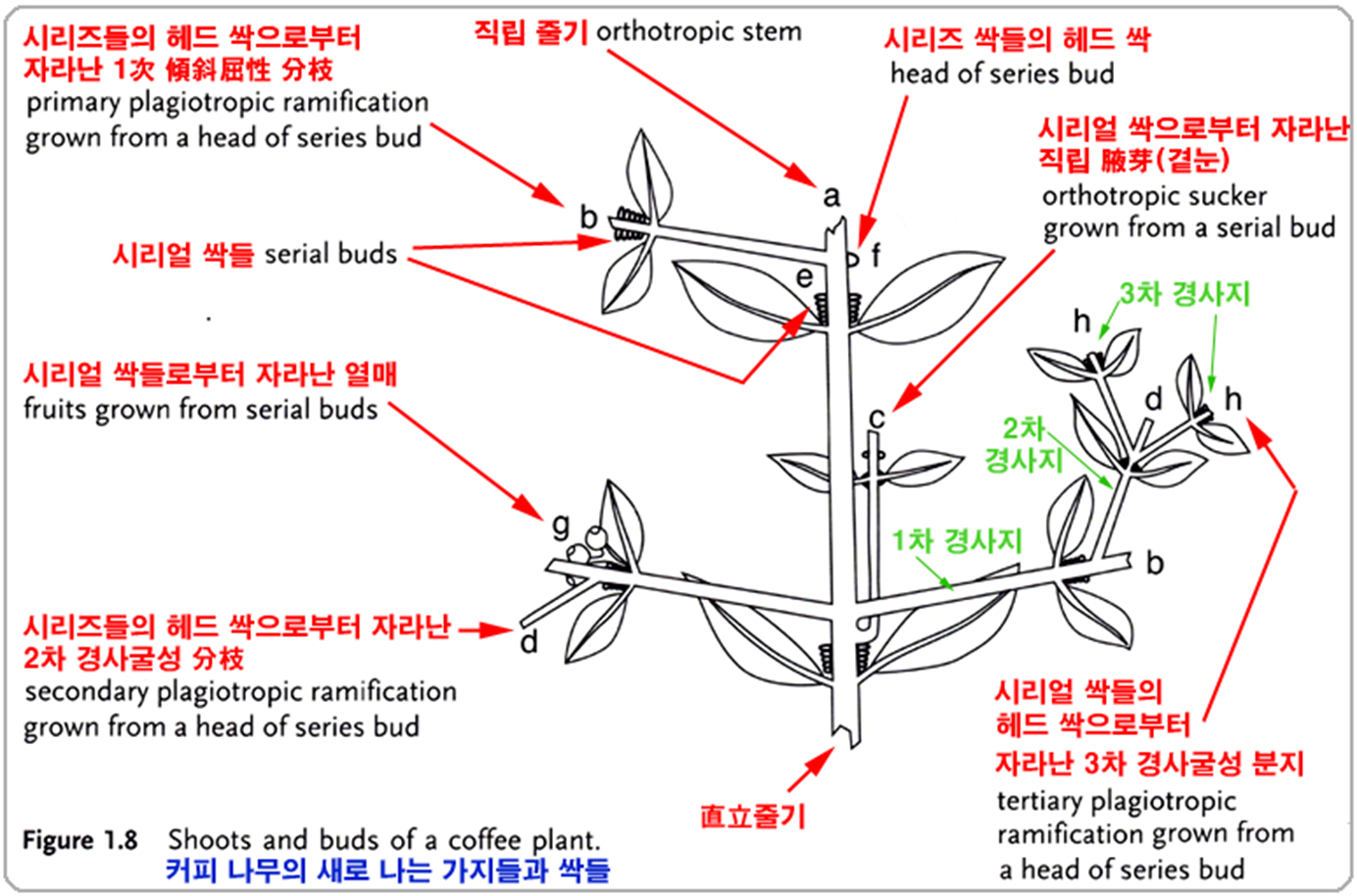
- 하나의 직립(single upright)(orthotropic, 직립성) 주 줄기(main stem)(트렁크, trunk)가 있으며,
⇒ 줄기의 지름은 나이와 성장 조건에 따라 다른데,
⇒ 나무 밑동에서 쟀을 때, 어린 나무의 경우 2~3cm, 오래된 나무의 경우 4~8cm 정도이다.
⇒ 매우 오래된 야생 나무들에서 줄기는 직경이 8~13cm이고], 드물게 15~20cm인 경우도 있다. - 몸통(trunk)에서 발생하는 주요 가지들(main branches)은
⇒ ‘側枝(laterals)’, ‘1次枝(primaries)' 또는 ‘一次枝梗(primary branches)'라고도 한다.
⇒ 주요 가지들은 주요 줄기(main stem) 위쪽으로
다소 일정하게 간격을 두고 마주보는 쌍(opposite pairs)으로 배열되며,
주 줄기와 직각으로(수평 또는 plagiotropic) 또는
낮은 각도(거의 수평, 최대 30° 또는 때로는 45° 이상)로 생긴다. - 一次枝梗은 반대 방향으로 배열되는 2次, 3次 및 때때로 4次 分枝를 발생시키며,
이는 종종 총칭하여 'sub-laterals(下位 側枝들)’라고 한다. - 나무가 pruned(剪枝, 가지치기), ‘topped'(꼭대기 잘림) 또는
‘stumped'(밑부분에서 잘림) 되거나, 또는 그 나무가 제대로 자라지 않으면,
모양이 바뀐다. - 또한, 10-20년 또는 그 이상이 되면 아라비카 나무는
⇒ 구부러지고(crooked)/또는 뒤틀린(twisted) 트렁크가 있는,
⇒ 어수선하고(untidy) 기울어진(leaning) 나무로 성장하여,
⇒ 반듯하고 다소 원추형인 수형과 규칙적인 가지 패턴을 잃는다. - 많은 나무가 수백 년, 심지어 천년 이상을 산다는 점을 고려하면,
아라비카 커피 나무는 수명이 짧은 것으로 분류된다. - 일반적으로 耕作되는 아라비카 나무는 10~15년 후에 최대 生産性을 멈추기 시작하며,
잘리지(stumped) 않거나 심하게 가지치기를 하지 않는 한, 총 20~30年 동안 산다. - 연간 나이테(tree ring) 數와, 주요 트렁크의 直徑(위 참조)과 같은 기타 증거를 보면,
매우 예외적인 환경에서 최대 100년, 어쩌면 140년까지 살 수 있음을 시사한다 (Tadesse, M. 2017). - 그러나 이런 더 오래 사는 아라비카 나무들은 여러 번 잘려 왔을 가능성도 높다.
- 野生 아라비카 나무들은 耕作되는 나무 보다 일반적으로 훨씬 더 느리게 자라며 더 오래 살 수 있다.

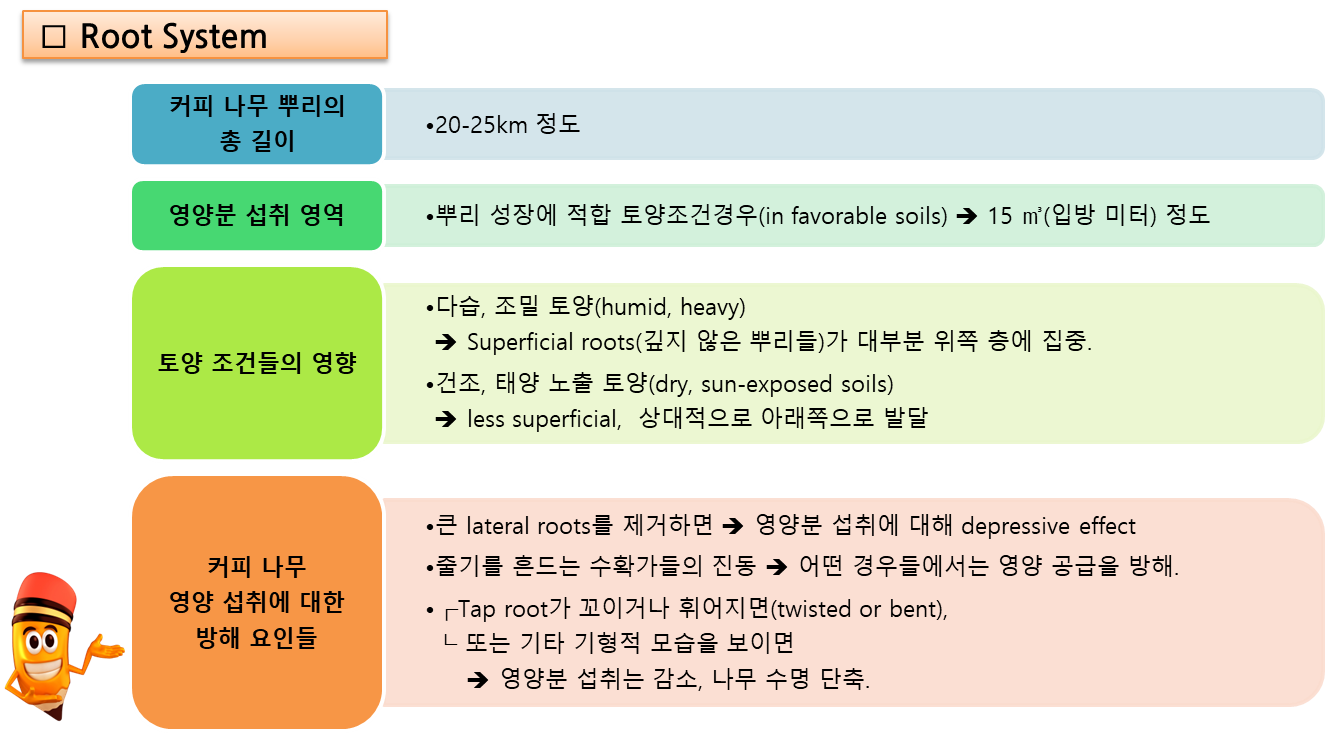
뿌리에 대한 다음 설명은 주로 커피나무 뿌리에 관한 고전적인 연구 작업인 Nutmann (1933a, 1933b, 1934)의 연구에 기반한다.
- 아라비카 커피 나무의 뿌리는 tap root (tap central root, 주근, 원뿌리), axial roots (직근, 축근), lateral roots (측근, 곁뿌리), (main) permanents (영구근), feeder bearers, 그리고 피더 (feeders, root hairs)로 구성된다.
- Tap root는, 보통 가지들을 내는데, 트렁크로부터 아래쪽으로 약 30~45cm 뻗어 있다.
- Axial roots는 tap root에서 4~8개가 나와서, 2~4m 아래로 땅속으로 자란다.
- lateral roots도 tap root에 붙어 있는데, 토양을 거쳐 다소 수평으로 자라서, 뿌리의 주 판(plate)을 형성하거나,
또는 아래쪽으로 달리고 여러 방향으로 가지를 뻗는다. - Permanents(영구근)는 axial roots와 lateral roots에서 발생하는 더 작고 목질인(woody)(cutinized) 뿌리이며(직경 약 3mm 또는 다소 더 넓음), 그 뿌리 시스템의 주요 골격(main framework)을 구성한다.
- Feeder bearers는 이것도 역시 목질이며(woody), permanents를 따라 다소 균등한 간격으로 자라며, feeders를 지탱한다.
- Feeders는 물과 영양분 흡수를 담당하며, 흰색이고 목질이 아니며(적어도 약 3년 동안) 대개 직경이 1.5mm 미만이다. 피더들 대부분은 보통 지표면에서 30~40cm 떨어져 있지만, 이는 많은 요인, 특히 토양 유형(soil type)과 물 가용성(water availability)에 따라 달라질 수 있다;
⇒ 비옥하고 깊은 토양(rich, deep soil)에서는 더 깊다.
⇒ 척박한 토양, 멀칭된 토양(천연적 또는 영농 실무를 통해), 그리고 관개 토양(irrigated soils)에서는
대부분 지표면에 더 가깝다 (Wrigley, G. 1988).



- 커피나무의 잎은
⇒ 반대로 배열(oppositely arranged),
⇒ 즉 잎자루(leaf stalks, 葉柄) (petioles)에 따라서 잎은 싹에 반대쪽 쌍으로 붙는다 (⇒ 對生葉). - 잎자루 사이에는 턱잎(stipules, 托葉)이라고 하는 구조 같은 작은 苞(bract),
또는 더 정확하게는 interpetiolar stiple(잎자루 사이의 턱잎)이라고 하는 것이 있으며,
모양은 다소 삼각형이다. - 잎은 상록수이지만 수명이 짧다. 각각의 잎사귀는 약 1년, 때로는 그 이상.
- 잎의 모든 부분에 털(hairs)이 없다 (즉, glabrous, 반들반들한 無毛이다).
- 잎의 크기(Leaf size)는
⇒ 수령, 환경, 유전적 기원에 따라 매우 다양하지만,
⇒ 보통 길이 7~18cm, 너비 3~9cm 사이이다. - 각 잎의 외곽선 모양(outline shape)은
⇒ 다소 타원형(가운데가 가장 넓고 끝으로 갈수록 가늘어짐)이며, - 잎 끝에는 작지만 뚜렷한 무딘 점(blunt point)(the acumen, 뾰족한 부분)이 있다.
- 잎의 둘레(margins)은 ⇒ 곧고 톱니가 있거나 들쭉날쭉하지 않다. 이러한 유형의 잎을 ‘simple'이라고 한다.
- 각 잎에는
⇒ 주맥(midrib) (main vein),
⇒ 6~10쌍의 2차 맥(secondary veins) (신경, nerves),
⇒ 수많은 미세한 3차 맥(finer tertiary veins)이 있다. - 잎은 처음에는 다소 얇으나 나이가 들면서 다소 가죽질(leathery) (subcoriaceous)이 된다.
- 잎의 윗면(upper surface)은
⇒ 짙은 녹색(dark green)이고 매끄럽고 윤기가 있으며(smooth and shiny), 매우 약간 밀랍(waxy); - 아래쪽 표면(lower surface)은
⇒ 밝은 녹색에서 중간 정도의 녹색이며 광택이 없다(matt, not shiny).
⇒ 잎의 밑면에는, 주맥과 2차맥 사이의 잎 겨드랑이(葉腋, axils)에는
도마티아(domatia)라고 하는 작고 부은 구덩이 모양의 구조가 있지만 육안으로는 보기 어렵다. - 두 가지 중요한 초기 커피 변종(variants)(종종 Coffea arabica의 varieties로 간주됨)인
'typica'와 'Bourbon'에 대한 주요 인식 특징 중 하나로
⇒ 어린 잎의 색상(colour of young leaves)이 두드러진다.
⇒ 'typica' 품종은 어린 잎과 잎 끝(leaf tips)이 청동색(bronze)인 반면
⇒ 'Bourbon' 품종은 녹색(green)이다.
⇒ 에티오피아의 야생 및 경작 아라비카 커피를 조사한 결과, 이 특성은 매우 가변적이며
두 유형 모두 가까운 곳에서 자라는 것을 볼 수 있다.
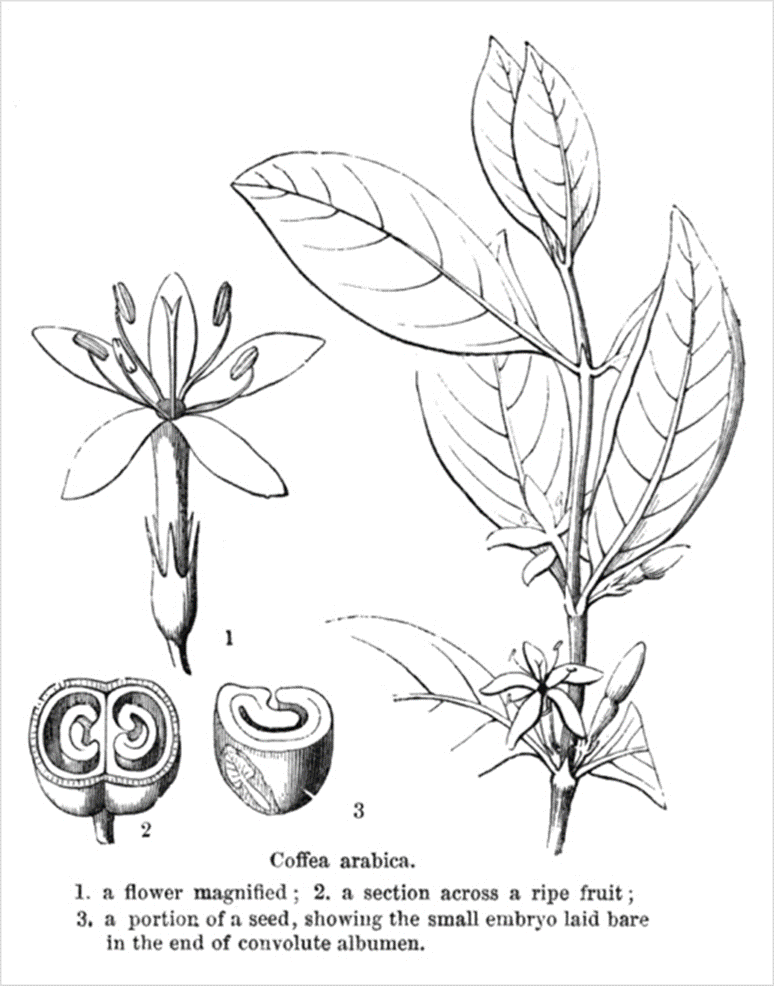


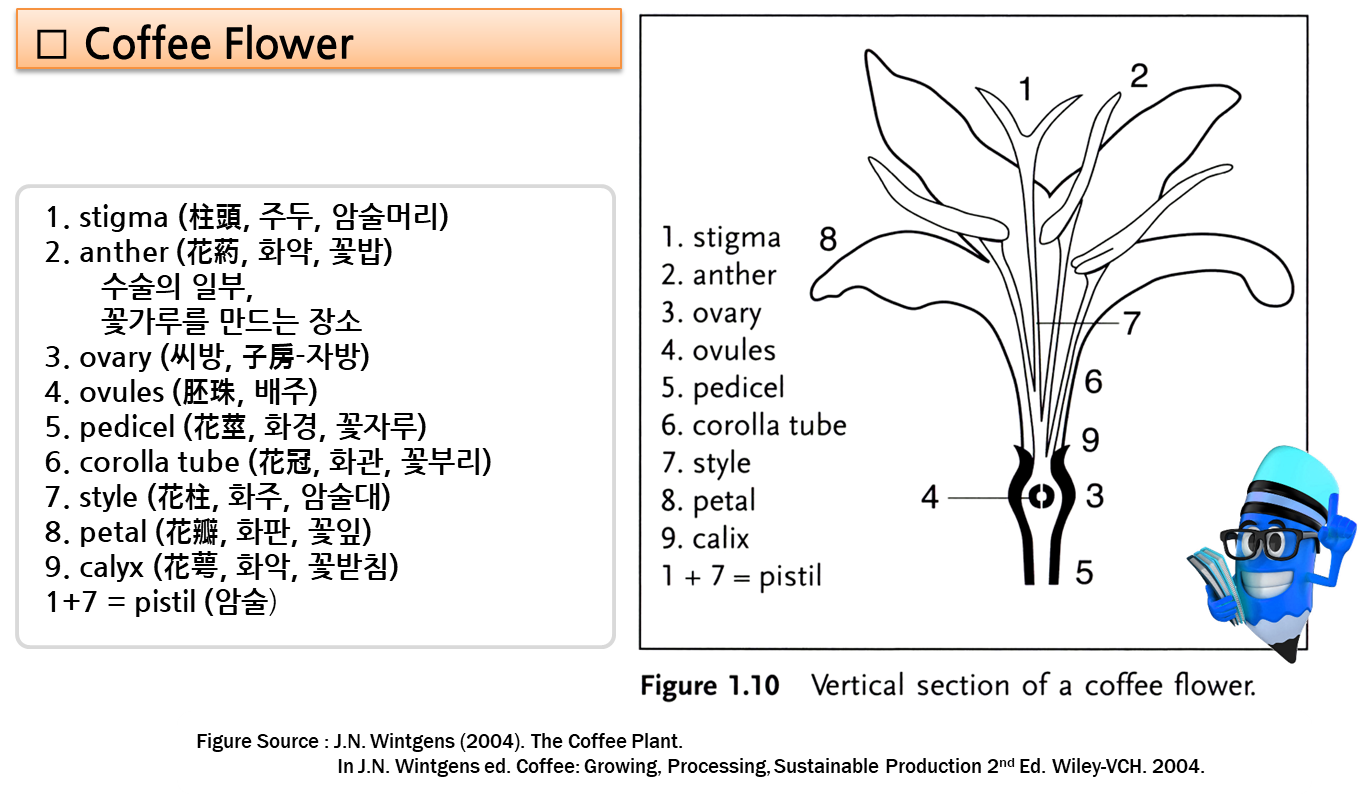
- 꽃은 ⇒ 잎의 겨드랑이(axils, 葉腋)에 위치한다.
⇒ 즉 (lateral)곁눈(shoot)과 잎의 자루(stalk)(petiole) 사이에 위치한다. - 각 잎 겨드랑이에는 1~10개의 꽃이 핀다.
즉, 한 송이(cluster)에(즉, 잎 쌍당)에 2~20개의 꽃이 있으며,
꽃차례(inflorescences)라고 하는 구조에서 작은 수로 함께 그룹화된다. - 숲이나 깊이 그늘진 곳에서 자라는 아라비카 커피 나무는 일반적으로 한 송이당 2개 정도의 적은 수의 꽃을 피운다.
- 각 곁눈(lateral shoot)은
⇒ 나무의 수령, 건강 및 유전적 특성에 따라 5-10개 또는 때로는 최대 12개의 꽃송이를 생산한다. - 잘 자라면 각 아라비카 커피 나무는 수만 송이의 꽃을 피울 것이다.
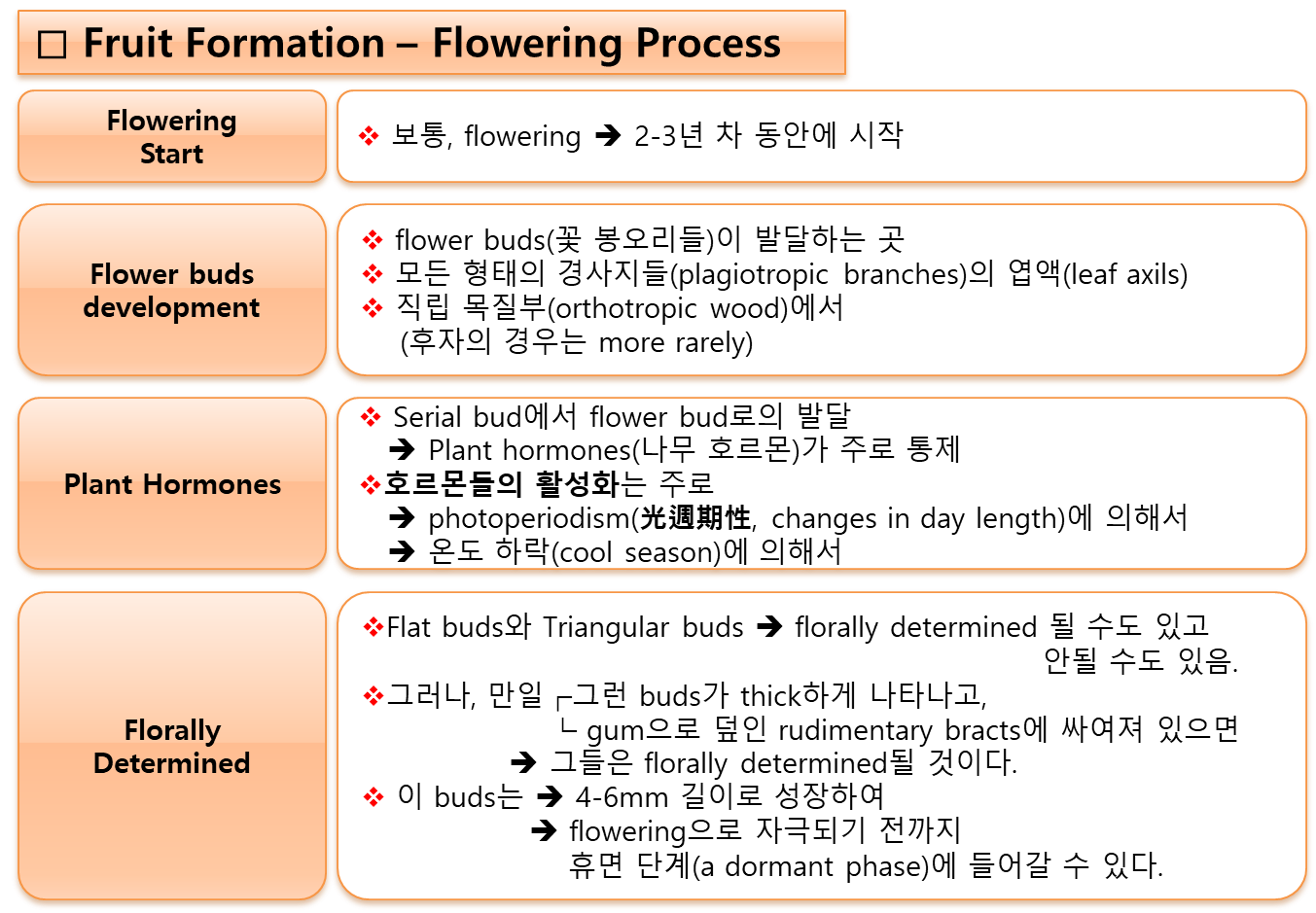
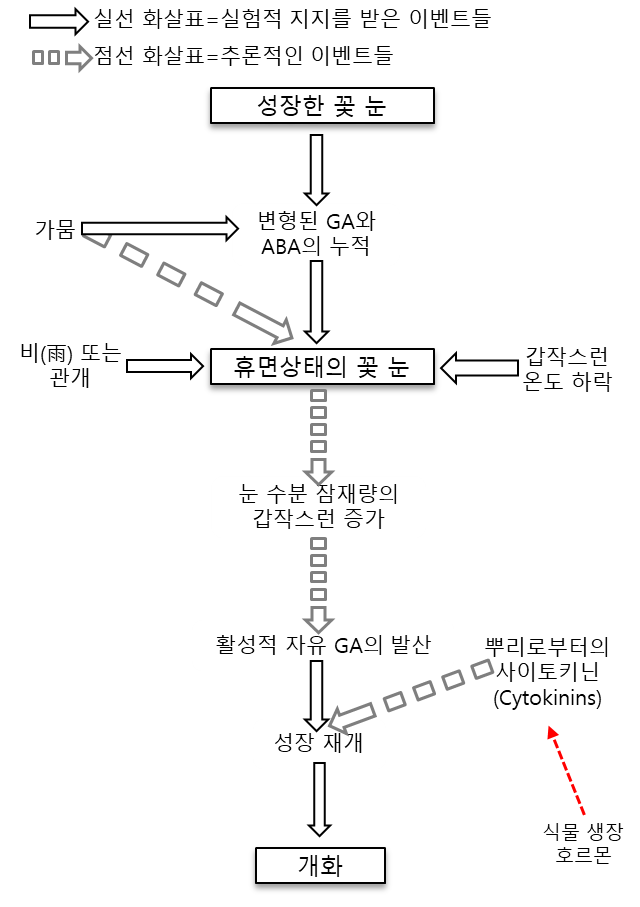
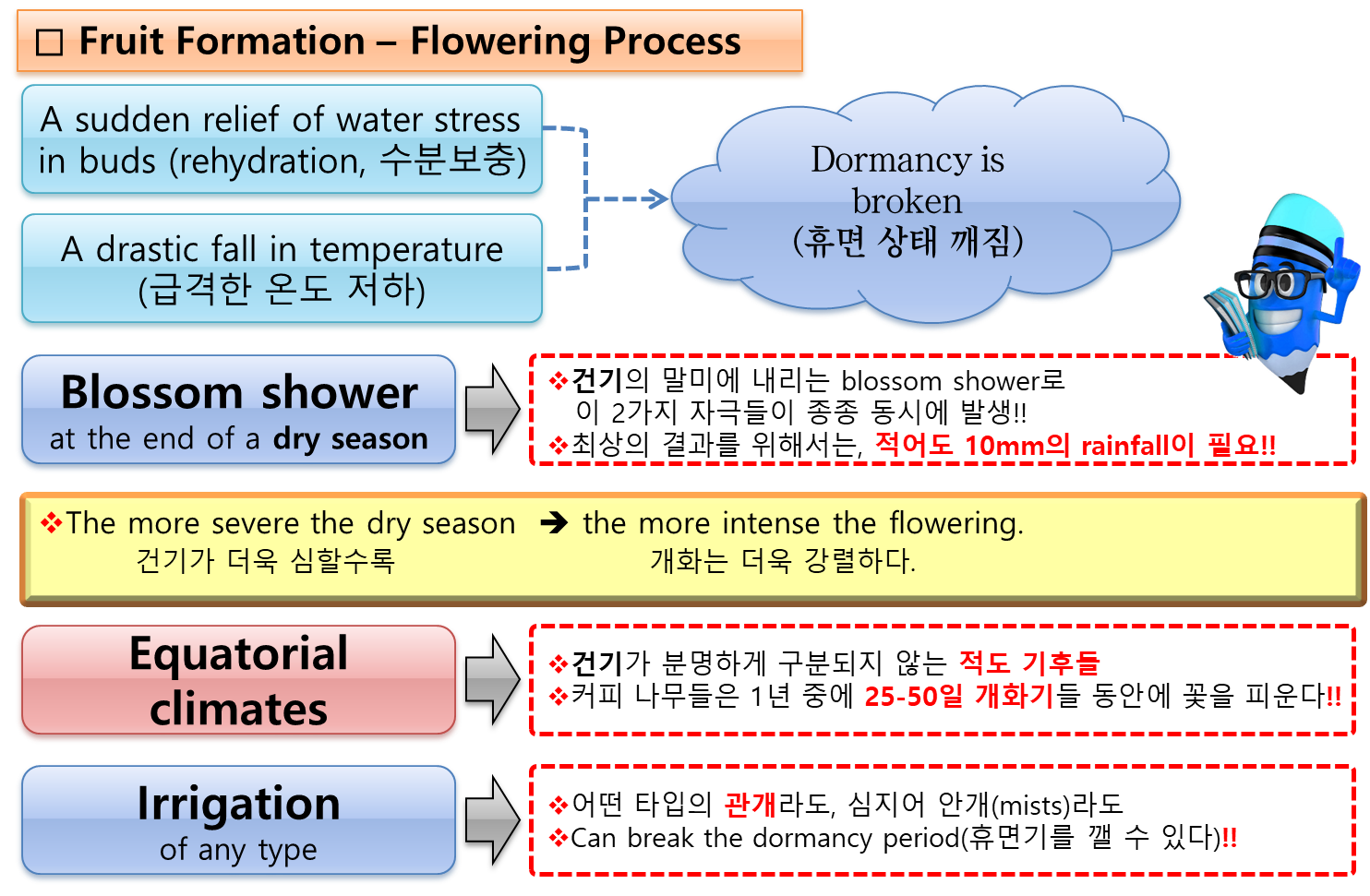
- 비가 오는 조건에서,
우기의 첫 번째 비로 인해 꽃봉오리(flower buds, 華蕾, 꽃눈)의 休眠狀態(dormancy)가 깨어지지만,
이는 3-4개월의 乾燥(休息) 其間이 선행되어야 한다. - 開化(flowering)는 보통 비가 온 후 8~12일 사이에 이루어지며, 온도 및 기타 요인에 따라 다르다.
- 꽃은 일반적으로 아침에 피고 정오 또는 오후에 시들어 곧 땅에 떨어지거나
며칠 동안 건조한 갈색 상태로 나무에 남아 있다. - 꽃의 주요 부분(corolla, 花冠, 꽃부리)은 흰색이며
基部(base)에서 결합하여 管 모양의 구조(corolla tube, 花冠筒部)를 형성하고,
그 위에 5개(때로는 4개 또는 6개)의 꽃잎(corolla lobes, 花冠 裂片)이 있다. - 화관 안에는 5개의 수술(stamens, 雄蘂)이 있다.
- 각 수술에는 꽃밥 주머니(anther sac, 花粉囊) (꽃가루가 들어 있는 구조)가 있는데,
꽃밥 주머니는 가느다란 줄기(fine stalk)(filament, 花絲)에 고정되어 있다. - 필라멘트의 다른 쪽 끝은 화관 내부로 부착되는데, 목 부위(화관의 최상부), 열편 사이 및 바로 아래에 붙는다.
- 수술들(stamens)은 흔히 '수컷’ 부분(male parts)이라고 한다.
- 꽃의 중앙에는 길고 가느다란 암술대(花柱, style)가 있으며,
끝 부위가 두 개의 암술대 편(style lobes)으로 끝나며, 꽃가루를 받는 (stigmatic) 표면(柱頭面)이 있다. - 基底部(base)에서 암술대는 디스크(disc)라고 하는 구조의 중심을 통과하여 씨방(ovary)에 연결된다.
- 각 씨방(子房)에는 각각 밑씨(胚珠, ovule)를 가지고 있는 두 개의 구획(compartments) (locules, 胞)이 있다.
- 꽃가루에 의해 授精되면 밑씨는 씨앗(커피콩)으로 발달한다.
- 암술대와 씨방은 종종 꽃의 '암컷’ 부분(‘female’ part)이라고 한다.

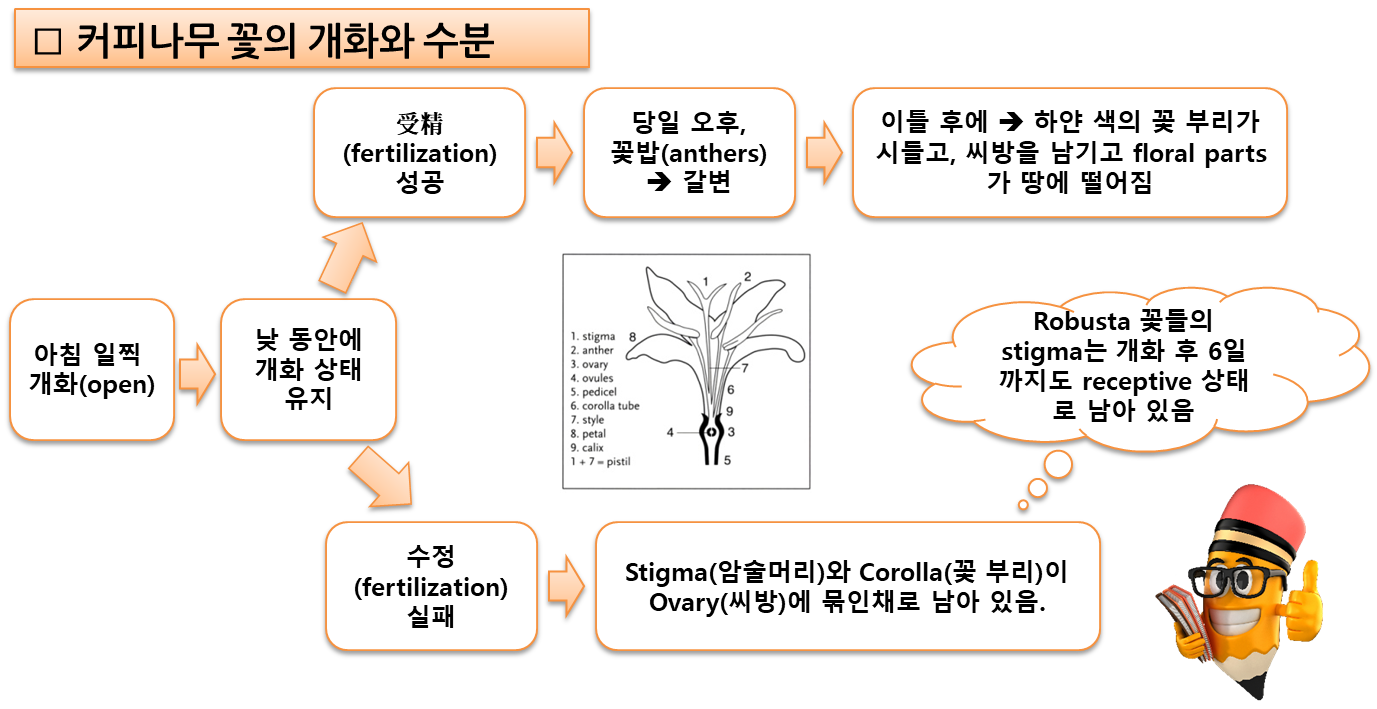

- 꽃은 앞서 설명된 대로
⇒ 꿀벌 등과 같은 곤충들에 의해 受粉 되거나(pollinated by bees)
⇒ 自家受精(self-fertilization)을 겪는다 (바람, 중력에 의한 수분). - 受粉 후 열매는
⇒ 처음에는 직경이 몇 밀리미터인 작은 녹색의 둥근 구조에서
⇒ 다양한 成熟 段階를 거쳐 성숙한 열매(식물학적으로 drupe(核果)로 알려짐)로 發達하기 시작한다. - 일반적으로 커피에서 일하는 사람들이 '체리'라고 부르는 잘 익은 과일은
⇒ 보통 빨간색이고 드물게 노란색이며
⇒ 바닥에 남아 있는 과일 줄기(fruit stalk)와
맨 위에 있는 원반(disc)(이전에는 꽃의 원반)을 제외하고는 부드럽다.
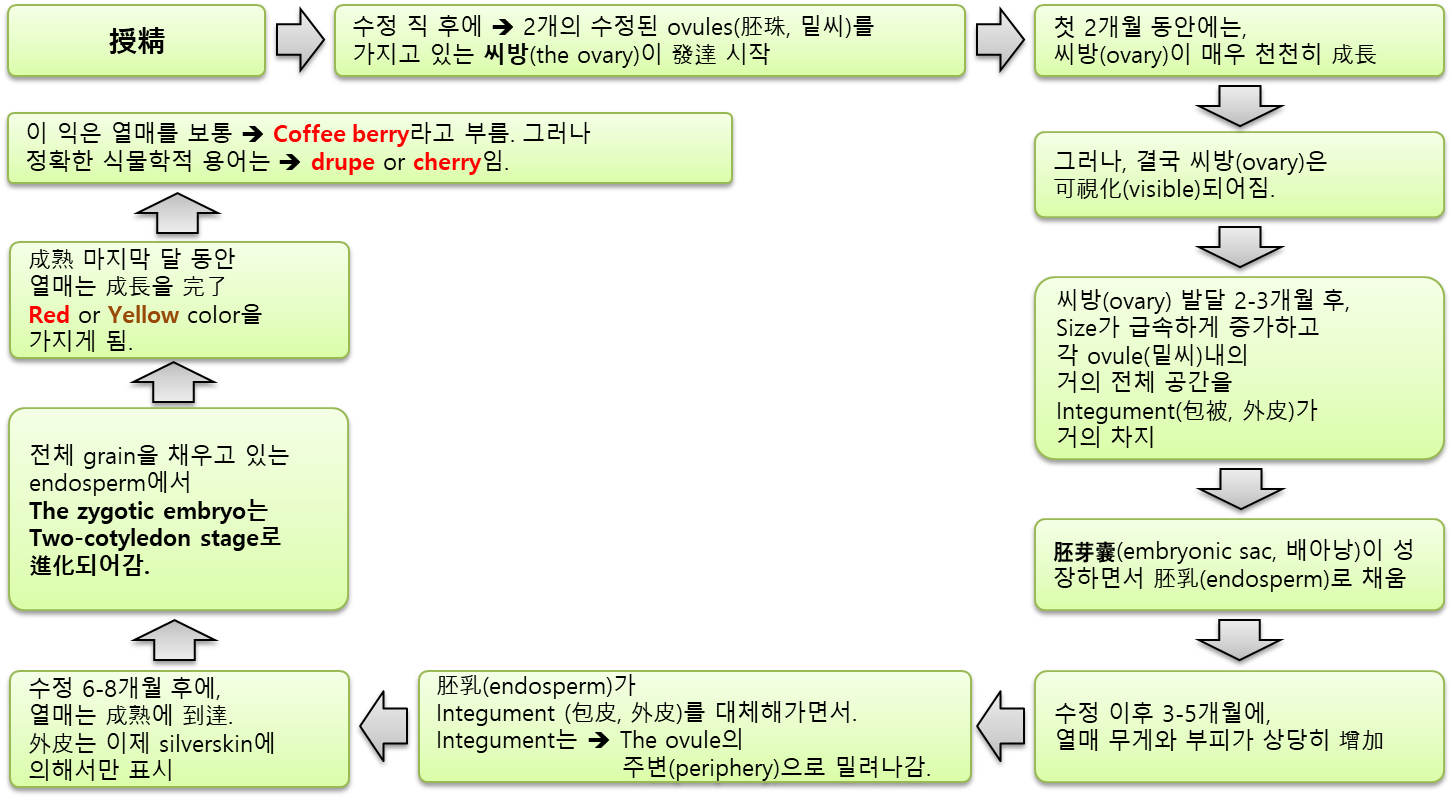
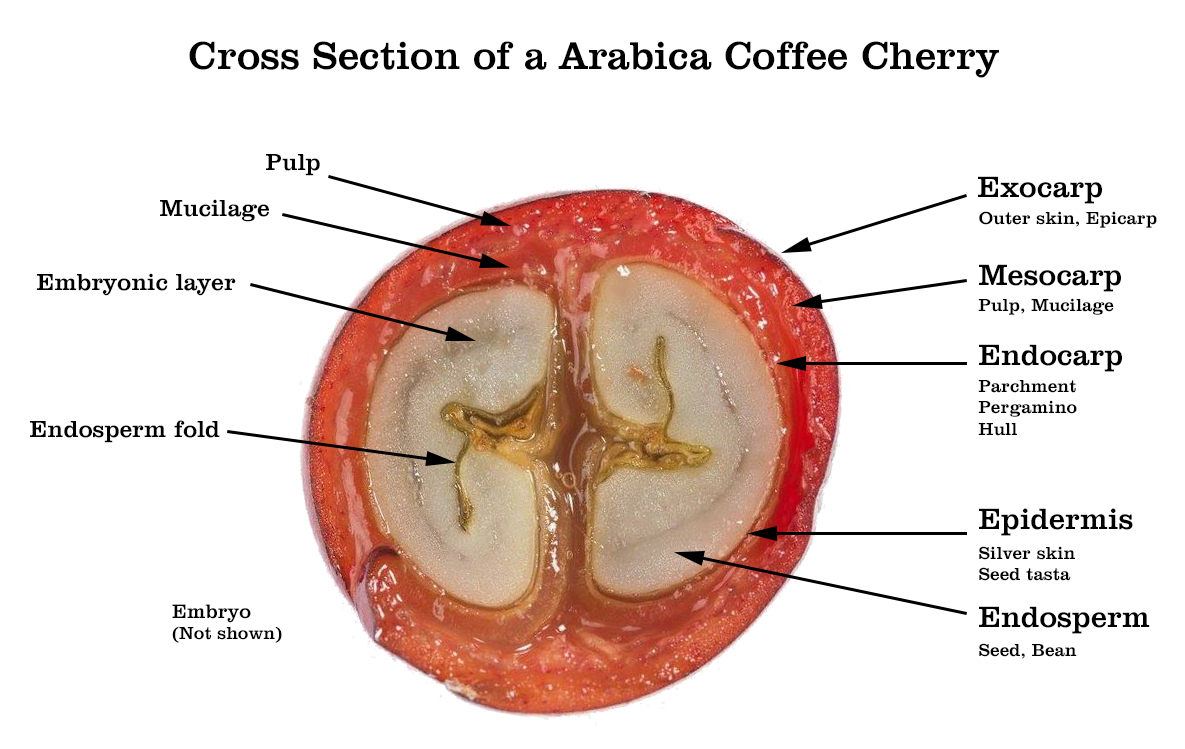
- 성숙한 열매는 여러 층으로 구성되어 있다.
- [outer skin] ⇒ 바깥쪽에는 열매 껍질(fruit skin) (epicarp, 外果皮)가 있고,
[pulp]⇒ 이 얇은 층 안에 수분이 많은 과육(watery pulp) (mesocarp, 中果皮)이 있으며,
[inner skin] ⇒ 그 다음에는 그린 커피 콩(seed)을 둘러싸는
파취먼트(parchment) 또는 hull (endocarp, 內果皮)이라고 하는
단단하고 바삭바삭한 층(내과피)이 있다.
[silver skin] ⇒ 씨앗은 얇은 細胞層인 실버 스킨(seed testa)으로 덮여 있는데,
말랐을 때는 거의 종이처럼 변한다.
[seed] ⇒ 씨앗의 내부에는 주로 탄수화물, 지질 및 단백질의 형태로
⇒ 작은 배아 식물(종자 기저부의 胚乳(endosperm) 내에 내장)에 영양을 제공하는
貯藏 細胞(storage cells) (endosperm, 內胚乳)로 주로 이루어진다. - 전체적으로 커피 열매는 익는 데 8~9개월이 걸린다.
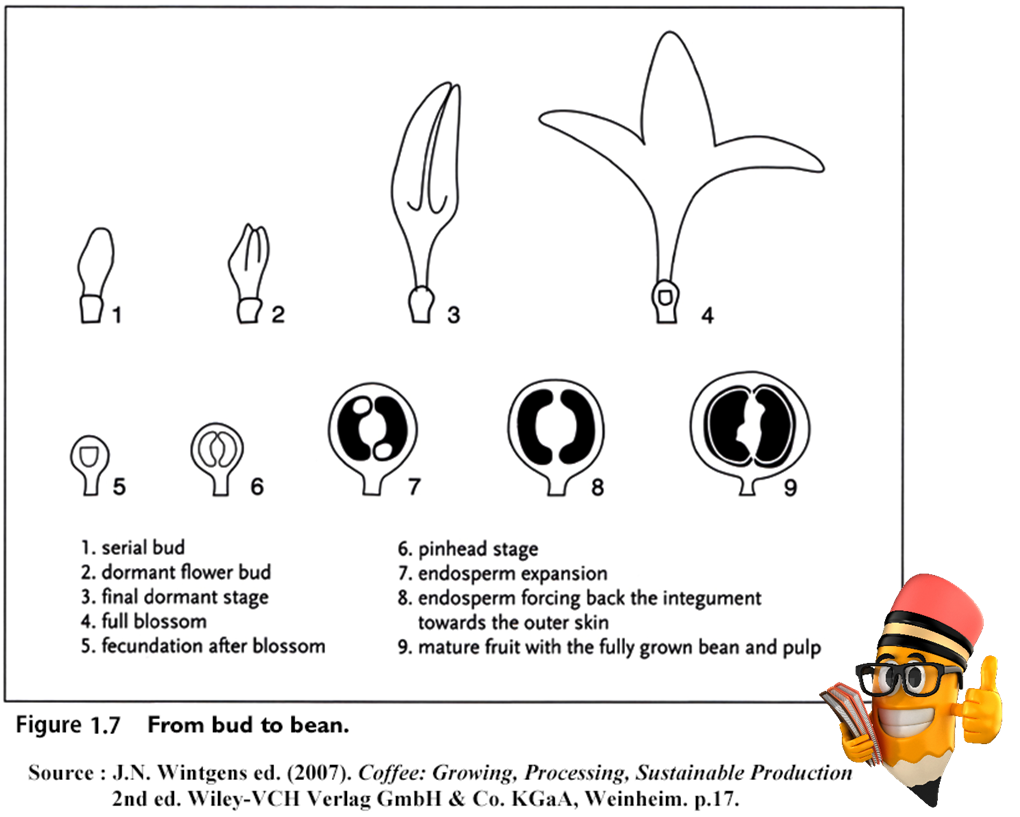
- 開花(및 受精) 후 처음 6~8주 동안 열매는 작고 둥글며 ‘pin-heads'라고 한다.
- 그 후, 8-18주부터 rapid growth phase(急速 成長段階)가 시작되는데,
이 때 과일의 내부 부분이 빠르게 발달하고 과일의 크기가 계속 커진다. - 그런 다음 약 2주 동안 지속되는 slow growth phase(느린 成長段階)가 있다 [30].
- 약 18주부터 32주까지 열매 발달(fruit development),
특히 배유(endosperm)의 발달과 건조 물질의 축적(accumulation of dry matter)이 계속된다. - 마지막으로 26주에서 36주 사이에
과일의 果肉 部分(pulpy part)(mesocarp, 中果皮)이 부풀어 오르고 물기가 많아지며,
열매 껍질(pericarp, 외과피)은 녹색에서 노란색으로 변하고 성숙기에 이르면 최종적으로 빨간색으로 변한다. - 發育速度(speed of development)는 降水量, 溫度, 土壤, 日照 時間 및 光度를 포함한 수많은 環境要因에 따라 달라진다.
- 일반적으로 果實 成熟段階(fruit ripening stage)는 약 10주 정도 걸리며
이 기간 동안 糖 含有率(sugar content)이 현저하게 增加한다.


- 天然 條件에서, 원숭이, 새, civets(사향 고양이)를 포함한 동물들이 숲과 숲 사이에 씨앗을 퍼뜨린다.
달콤하고 영양가 있는 과일은 動物이 섭취하는데, 이 과정에서 과일의 果肉 部分을 모두 제거하고 씨앗은 그대로 둔다.
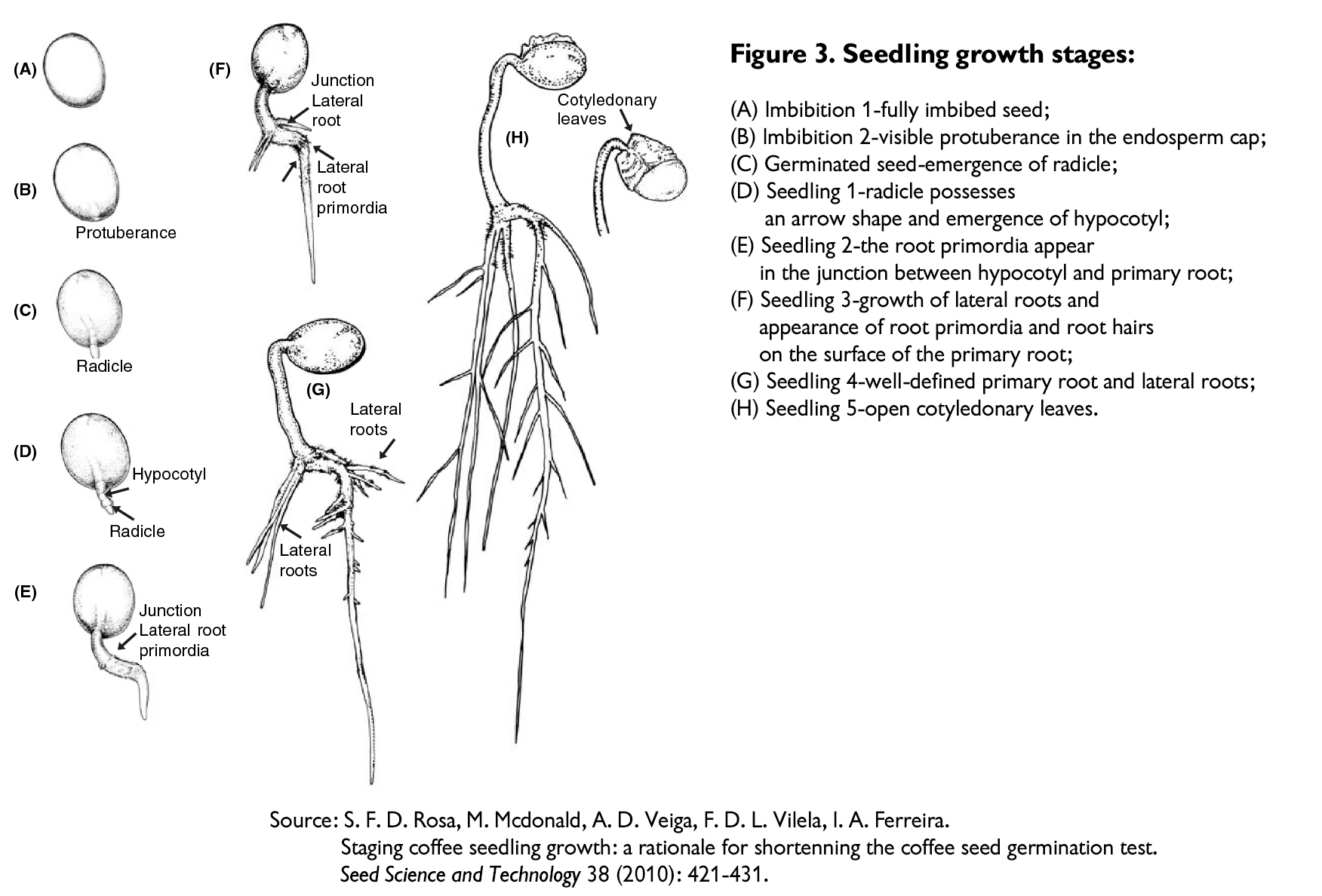
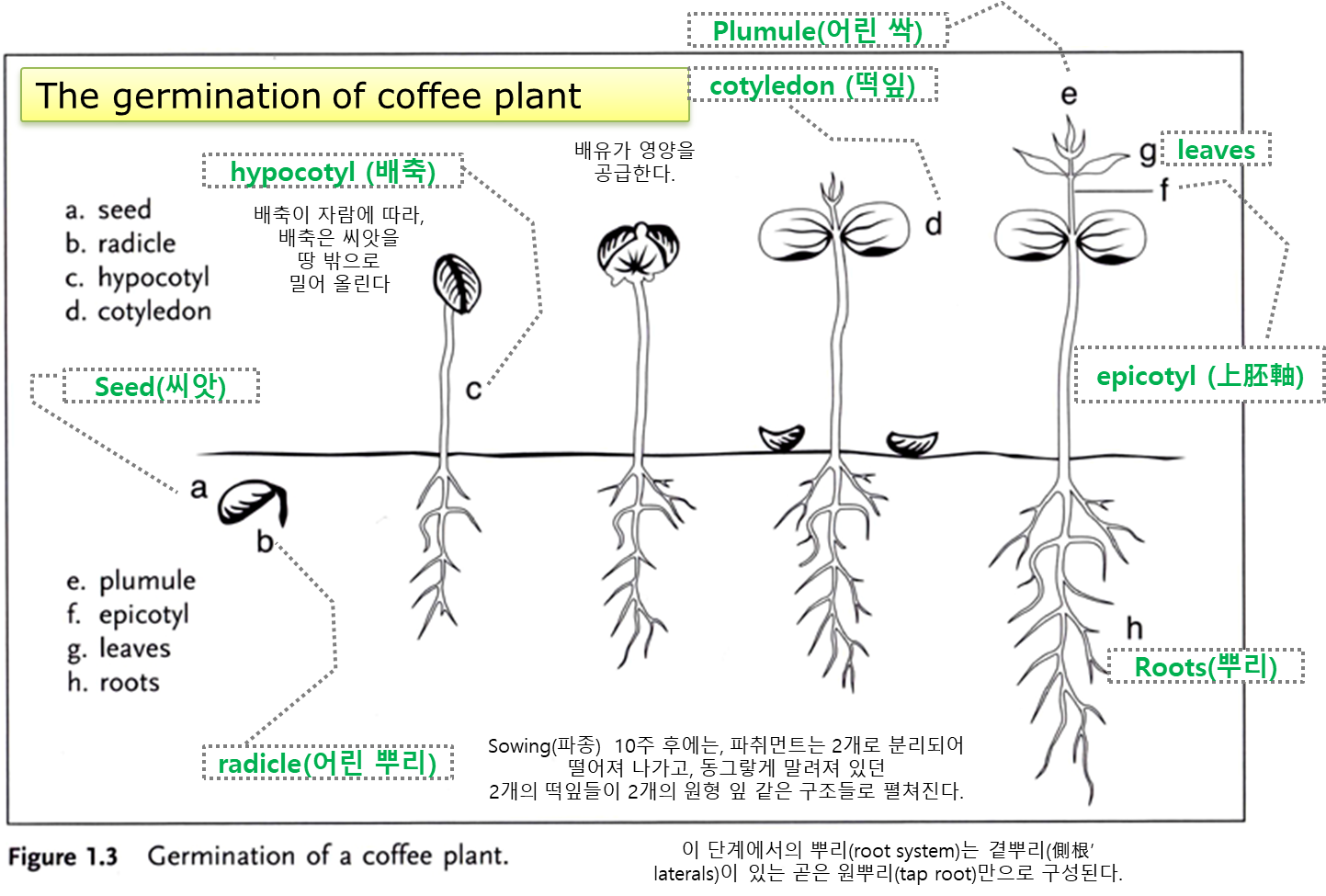
씨앗은 동물의 배설물 속에서 땅으로 배설되며, 비를 맞고 發芽할 준비가 된다. - 發芽(germination) 직후 씨앗은 어린 줄기(young stem)의 흙 위로 밀려나오는데,
이 단계에서 씨앗은 'soldier'라고 불린다. - 땅 위의 씨앗 내부에는 처음 두 개의 어린 잎(cotyledons, 子葉, 떡잎)이 있는데,
이 잎은 배유로부터 備蓄 物質을 계속 끌어내고
결국(3~4주 후) 남은 씨앗(대부분 파취먼트의 잔해)을 밀어낸다. - 子葉은 대략 귀 모양이므로, 일반 잎과 매우 다르게 보인다.
- 얼마 지나지 않아 진정한 잎들이 나타나고 더 많은 잎 쌍이 발달한다.
- 이 단계에서부터 그 어린 나무는 자라고 成長하여 成熟하게 된다.
- 栽培의 경우에서는, 成熟(maturity)은 첫 작물(first crop)이 열릴 때로 정의되며,
발아 후 약 4~5년이 걸린다. - 野生에서는, 성장(growth)은 주로 다른 식물과의 경쟁으로 인해 일반적으로 훨씬 더 느리다.
결실 단계는 발아 후 10년 또는 그 이상이 될 때까지 도달하지 못할 수 있다.
| References |
- Friis, I., Demissew, S. & Breugel, P. V. (2010). Atlas of the potential vegetation of Ethiopia.
Biol. Skrif. 58: 1–307 - Davis, A. P., Govaerts, R., Bridson, D. M. & Stoffelen, P. (2006).
An annotated taxonomic conspectus of the genus Coffea (Rubiaceae).
Botanical Journal of the Linnean Society, volume 152, issue 4 (2006), pp. 465–512.
DOI: 10.1111/j.1095-8339.2006.00584.x. - Tesfaye, K., Borsch, T., Govers, K. & Bekele, E. (2007).
Characterisation of Coffea chloroplast microsatellites and evidence for the recent divergence of C. arabica and C. eugneioides chloroplast genomes. Genome 50: 1112–1129. - Davis, A. P., Gole, T. W., Baena, S. & Moat, J. (2012).
The impact of climate change on natural populations of Arabica coffee: predicting future trends and identifying priorities. PLoS ONE 7: e47981. - Davis, A. P., Tosh, J., Ruch, N. & Fay, M. (2011). Growing coffee: Psilanthus (Rubiaceae) subsumed on the basis of molecular and morphological data; implications for the size, morphology, distribution and evolutionary history of Coffea. Bot. J. Linn. Soc. 167: 357–377.
- Senbeta, F., Denich, M., Böhmer, H. J., Gole, T. W., Teketay, D. & Demissew, S. (2007).
Wild Coffea arabica L. in the Afromontane rainforests of Ethiopia: distribution, ecology and conservation.
SINET: Ethiop. J. Sci. 30: 13–24. - Mittermeier, R. A., Robles-Gil, P., Hoffmann, M., Pilgrim, J., Brooks, T., Mittermeier, C. G., Lamoreux, J. & Da Fonseca, G. A. B. (2004).
Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions.
CEMEX (Mexico City). - Meyer, F. G. (1965).
Notes on wild Coffea arabica from southwestern Ethiopia, with some historical considerations.
Econ. Bot. 19: 136–151. - Tadesse, M. (2017). Ethiopia — Home of Arabica Coffee: Early Use, Folklore, Coffee Ceremony, Origin and Biology.
CreateSpace Independent Publishing Platform (North Charleston, South Carolina). . - Sylvain, P. G. (1958). Ethiopian coffee: its significance to world coffee problems. Econ. Bot. 12: 111–139.
- Nutman, F. J. (1933a). The roots system of Coffea arabica L. I: Root systems in typical soils of British East Africa.
Empire J. Exp. Agric. 1: 271–284. - Nutman, F. J. (1933b). The roots system of Coffea arabica L. II: The effects of some soil conditions in modifying the ‘normal’ root system. Empire J. Exp. Agric. 1: 285–296.
- Nutman, F. J. (1934). The roots system of Coffea arabica L. III: The spatial distribution of the absorbing area of the root. Empire J. Exp. Agric. 2: 293–302.
- DaMatta, F. M., Ronchi, C. P., Maestri, M. & Barros, R. S. (2007). Ecophysiology of coffee growth and production.
Braz. J. Plant Physiol. 19: 485–510. - Hailu, B., Maeda, E. E., Heiskanen, J. & Pellikka, P. (2015).
Reconstructing pre-agricultural expansion vegetation cover of Ethiopia. Appl. Geogr. 62: 357–365. - Wrigley, G. (1988). Coffee — Tropical Agriculture Series. Longman Scientific & Technical, Harlow, UK. 639 pages.
- J.N. Wintgens ed. (2007). Coffee: Growing, Processing, Sustainable Production 2nd ed.
Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim. - Justin Moat, Jenny Williams, Susana Baena, Timothy Wilkinson, Tadesse W. Gole, Zeleke K. Challa, Sebsebe Demissew, and Aaron P. Davis (2017).
Resilience potential of the Ethiopian coffee sector under climate change. NATURE PLANTS 3, 17081 (2017).
DOI: 10.1038/nplants.2017.81 - Aaron P. Davis, Catherine Kiwuka, Aisyah Faruk, John Mulumba and James Kalema.
A review of the indigenous coffee resources of Uganda and their potential
for coffee sector sustainability and development. Front. Plant Sci., 17 February 2023.
DOI: 10.3389/fpls.2022.1057317

- The end -
'Ethiopia Coffee' 카테고리의 다른 글
| 에티오피아 커피 [6] 커피경작시스템 (0) | 2023.05.31 |
|---|---|
| 에티오피아 커피 [5] 커피 기후와 농생태학 (0) | 2023.05.26 |
| 에티오피아 커피 [3] 커피 세리모니 및 의례 (0) | 2023.05.20 |
| 에티오피아 커피 [2] 커피 소비 (0) | 2023.05.10 |
| 에티오피아 커피 [1] 개요 (1) | 2023.05.08 |




댓글