
Contents
| ■ Abstract | ||
| ■ Introduction | ||
| ■ Chemosensory processes in the tongue | ||
| Taste bud cell types and taste perception | ||
| Type I cells | ||
| Type II cells | ||
| Type III cells | ||
| Taste cell precursors | ||
| Signalling mechanisms of taste perception |
||
| Hormones expressed in taste buds | ||
| CCK and Y‑family peptides | ||
| Glucagon, GLP‑1 and GLP-2 |
||
| VIP peptides |
||
| Ghrelin | ||
| Oxytocin | ||
| Galanin | ||
| Leptin | ||
| TBCs have similarities to islets of Langerhans |
||
| ■ The gut as a chemosensory organ |
||
| Sweet taste receptors |
||
| Bitter taste receptors | ||
| Fat-sensing receptors | ||
| Endocannabinoid signalling | ||
| Role of the gut microbiota in food sensing | ||
| ■ Chemosensory processes in the pancreas | ||
| ■ Neuronal control of taste | ||
| ■ Taste perception and obesity | ||
| Surgical interventions affecting taste and food intake | ||
| Weight-loss surgery |
||
| Vagotomy | ||
| ■ Conclusions | ||
| ■ References | ||
Abstract
- 肥滿은 전 세계적으로 유행병(epidemic) 수준에 도달하여 건강 문제가 상당히 증가하고 심혈관 질환(cardiovascular diseases), 폐 질환(pulmonary diseases), 암(cancer), 당뇨병(diabetes mellitus)을 포함한 여러 질병의 위험도 증가했다.
- 비만 환자들은 과체중에 따른 차별과 과식이 건강에 해롭다는 풍부한 정보에도 불구하고 이상적인 체중을 유지하는 데 필요한 것보다 더 많은 음식을 섭취한다.
- 개인의 평생 에너지 소비와 섭취 사이의 관계는 이전에 생각했던 것보다 훨씬 더 복잡하다.
- 맛(taste), 기호성(palatability, 입에 맞음), 미각 수용체(taste receptors), 그리고 음식에 대한 쾌락 반응(hedonic responses) 사이의 관계에 대한 이해가 향상되면, 에너지 획득의 생물학적 기저에 대한 이해는 물론, 인간이 때때로 필요 이상으로, 그리고 건강에 좋다고 생각하는 것보다 더 많이 섭취하는 이유에 대한 이해도 높아질 것이다.
- 본고에서는
혀(tongue), 장(gut), 췌장(pancreas), 뇌(brain)에 있는 미각 수용체의 역할과,
맛 지각 (taste perception)에 대한 그들 호르몬의 관여, 그리고 맛 지각(taste perception), 과식(overeating), 그리고 비만 발생(development of obesity) 사이의 관계를 논의한다.
Key points
- 과식이 해롭다는 것을 알면서도 과체중인 많은 사람들은 음식 섭취를 조절하지 못한다.
- 식사를 통해 얻는 만족감, 즉 쾌락 반응은 포만 피드백 메커니즘(satiety feedback mechanisms)을 극복한다.
- 혀의 味細胞(taste cells)에서 생성되는 호르몬은 맛 지각의 강도(intensity of taste perception)를 변화시킨다;
렙틴(leptin)은 먹는 것(eating)에 대한 신경학적 쾌락 반응(neurological hedonic responses)과 단맛 지각의 강도(intensity of sweet perception)를 변화시킨다. - 미각 수용체(taste receptors)의 부위는 미세포에만 국한되지 않으며, 다른 생리 기능에서 이러한 수용체의 역할이 연구되고 있다.
- 비만(Obesity) 및/또는 과영양(overnutrition)(만성적 과잉 에너지 상태)은 맛 지각에 영향을 미칠 수 있다;
비만인 사람은 건강한 사람과 동일한 강도의 쾌락 반응을 유발하기 위해 더 많은 량의 맛 물질(tastants)이 필요하다. - 대사 수술(metabolic surgery)이 체중 감량에 미치는 영향과 맛 지각에서 장내 미생물군(gut microbiota)의 역할에 대한 이해는 맛 지각과 비만의 관련성을 밝혀낼 수 있다.
Introduction
칼로리가 높은 음식을 쉽게 구할 수 있고 칼로리를 얻기 위해 거의 또는 전혀 노력할 필요가 없는 사회에서 많은 개인은 건강한 체중을 유지하기 위해 음식 섭취를 조절하는 데 어려움을 겪는다. 비만 위험과 관련된 유전자와 음식 섭취, 포만감, 음식 소화 및 음식 흡수의 내분비학에 관련된 유전자를 식별하는 데 큰 진전이 있었다 [1]. 그러나 비만(obesity)과 당뇨병 유행(diabetes epidemics)은 지속되고 가속화되는 것으로 보인다 [2]. 질병 통제 예방 센터에서 발표한 보고서에 따르면 2012년 현재 미국에는 진성 당뇨병(diabetes mellitus) 환자가 2,900만 명, 당뇨병 전단계(pre-diabetes) 환자가 8,600만 명이다. 영향을 받는 환자의 대다수는 비만 또는 과체중과 관련된 2형 당뇨병(type 2 diabetes mellitus)을 앓고 있다 [3].
포유류의 생리 시스템(Mammalian physiological systems)은 기분을 좋게 하려는 동물의 욕구가 가장 중요하도록 발달해왔다 [4]. 맛있는 음식(Palatable foods)에는 지방(fat), 단맛을 내는 화학 물질, 약간의 소금(salt)이 들어 있다; 이런 음식을 쉽게 구할 수 있다면, 맛이 좋고 기분도 좋아지기 때문에 과도하게 먹게 된다 [5]. 포만감(satiety)을 결정하는 장(gut)의 분자 신호는 지각이 있는(sentient) 인간이 쉽게 조작할 수 있으며, 체지방 함량을 나타내는 비만 신호와 시각적 단서도 마찬가지이다 [4,6,7]. 예를 들어, 개인이 과체중에서 비만으로 진행됨에 따라, 렙틴 (leptin, 전신 지방과다(adiposity)의 호르몬 지표)의 순환 수치는 증가하지만, 렙틴 수용체의 발현은 하향 조절된다 [8,9].
일반적으로 glucagon-like peptide 1 (GLP‑1, 글루카곤-유사 펩타이드 1) 및 cholecystokinin (CCK, 콜레시스토키닌)과 같이 포만감을 조절하는 장 호르몬은 빠르게 분해되기 때문에 반감기가 매우 짧다. 따라서 이러한 호르몬이 배고픔과 포만감에 미치는 영향은 음식 섭취 사이에 짧은 간격을 두고 자주 먹음으로써 조절할 수 있다 [1]. 맛있고 가공된 값싼 음식, 특히 설탕이 함유된 음식에 반복적으로 노출되면 음식에 대한 즐거움이 강화되어 과식으로 이어진다 [10].
호르몬이 미각 반응을 어떻게 조절하는지에 대한 포괄적인 이해를 위해서는, 맛 지각(taste perception)의 기저에 있는 해부학적, 생리학적, 생화학적 구조를 이해하는 것이 필수적이다. 뛰어난 연구들을 통해 미각 반응 형성에 있어 고전적 및 비전통적 소분자 신경전달물질(neurotransmitters)의 역할에 대한 정보가 도출되었으며, 이 주제는 다른 논문에서 광범위하게 검토되었다 [11–13]. 본 리뷰에서는, 맛 지각의 기저에 있는 생물학적 메커니즘, 미뢰에서 생성되는 호르몬과 그 호르몬의 맛 지각에 대한 관여, 그리고 장(gut)과 췌장(pancreas)의 미각 수용체에 대해 논의한다. 또한, 당 섭취에 대한 신경 반응도 다룬다.
Chemosensory processes in the tongue
혀에서의 화학감각적 프로세스
Taste bud cell types and taste perception
미뢰 세포 유형과 맛 지각
Taste (맛), 또는 gustation (미각)은 5가지 주요 감각들 중 하나이다. 맛 지각(taste perception)은 화학 물질이 혀의 미뢰 세포(taste bud cells, TBC)와 접촉할 때 촉발된다; TBCs의 아세포들(하위집단)은 장과 뇌에서도 발견되는 호르몬(예: GLP-1 및 ghrelin 그렐린)을 합성한다 [14,15].
대부분의 동물은 다섯 가지 기본 맛(basic tastes)(단맛, 감칠맛(우마미), 쓴맛, 짠맛, 신맛)을 인지하며, 지방도 느낄 수 있다는 증거가 점차 늘어나고 있다 [16,17].
미뢰는 마늘과 유사한 구조를 가지고 있으며(Figure 1a), 주로 혀의
■ 有廓乳頭(circumvallate papillae),
■ 葉狀乳頭 (foliate papillae),
■ 菌狀乳頭 (fungiform papillae, 茸狀乳頭)에 위치한
미각유두(taste papillae) 내에 존재한다 (Figure 1b).
일부 분리된 미뢰들과 G-protein-coupled taste receptors (taste GPCR)를 발현하는 화학감각 세포(chemosensory cells)는 쥐(rats)의 口蓋(palate), 喉頭蓋(epiglottis), 咽頭(pharynx), 喉頭(larynx) 및 鼻咽頭切齒管(nasoincisor duct, 코앞니관)에 존재한다 [18,19].
각 미뢰에는 마늘과 같은 구조의 ' cloves (마늘 쪽들)'로 조립된 약 50~100개의 TBCs가 들어 있으며, 신경성 전구체(neurogenic precursors)에서 발생하는 후각 수용체 세포(olfactory receptor cells)와는 달리 상피 세포(epithelial)에서 유래한다 [20].
TBC는
⇒ 4가지 하위 타입들로 분류되며
⇒ 모든 미뢰에는 해부학적 위치와 관계없이 4가지 하위 타입의 세포가 모두 들어 있다 (Figure 1a).

▣ Figure 1 | 인간 혀(tongue)에서의 미뢰(taste bud) 위치 및 구조.
a | 미뢰와 미뢰 내 신경 섬유(intragemmal nerve fibres)의 도해적 표현.
일반적으로 미뢰에는 네 가지 서브타입의 미뢰 세포가 존재한다.
네 가지 서브타입 중 type III 미뢰 세포만이 求心性 神經纖維와 인지 가능한 시냅스를 형성한다.
b | 미각 유두(taste papillae)의 위치.
Circumvallate (遊廓乳頭)(혀의 뒤쪽),
foliate (葉狀乳頭)(혀의 측면), 그리고
fungiform(菌狀乳頭)(혀의 중간과 앞쪽)에 위치한다.
Type I cells
- TBC의 총 수의 약 50%는 미뢰의 지지 구조를 유지하는 type I cells이다.
- 이 세포는 뚜렷한 전기 생리학적 특징이 있다.
- Type I TBC는
⇒ 작은 voltage-gated outward K+ and inward Na2+ currents을 가지지만,
⇒ 전압 개폐성 Ca2+ currents는 없다 [21]. - 아밀로라이드-민감성 나트륨 채널 서브유닛 α (Amiloride-sensitive sodium channel subunit α)
(일반적으로 epithelial sodium channel subunit α [α ENaC]라고 함)가
⇒ type I cells에서 발현되며
⇒ 저염(예: NaCl 또는 KCl) 지각의 주요 조정체(mediator)로 간주된다 [22,23]. - 마우스에서 TBC-특이적 α‑ENaC 삭제로 인해 소금에 대한 행동적 끌림이 완전히 상실되었다 [23]; 그러나 저염 맛 물질이 type I cells를 활성화할 때 활성화되는 하류 신호 전달 메커니즘과 이 세포가 신경 섬유와 통신하는 방법은 알려져 있지 않다.
- 이 세포들은 α‑ENaC를 발현하는 것 외에도,
세포 표면에 membrane-bound ATPase를 발현하여 이웃 세포에서 방출된 ATP를 분해한다. - Type I cells는
⇒ 미뢰 구조 내에서 다른 세포 타입들을 둘러 싸는
넓은 판상 돌기(층상 돌기, lamellar processes)를 가지고 있으며,
⇒ 이는 미뢰 전체에서 세포 신호 전달 분자의 소멸(dissipation)을 컨트롤하고
미뢰의 특정 영역으로의 이온 변동(ion fluctuations)을 분리하는 기능을
하는 것으로 추정된다 [24, 25].
Type II cells
- Type II cells는
⇒ 종종 receptor cells (수용체 세포)라고도 하며,
⇒ 단맛(sweet), 우마미(umami), 쓴맛(bitter) 물질들을
감지하는 수용체들을 발현한다 [26–28]. - 단맛과 우마미 맛 물질은
⇒ 세 가지 수용체(TAS1Rs)로 구성된 이종이량체(heterodimeric) GPCR에 의해 감지된다:
⇒ 이 수용체들은
■ taste receptor type 1 member 1 (TAS1R1),
■ taste receptor type 1 member 2 (TAS1R2) 그리고
■ taste receptor type 1 member 3 (TAS1R3)이다. - TAS1R1 및 TAS1R3 서브유닛의 이종이량체 수용체는 우마미 미각 물질(예: 글루탐산, 국물, 버섯, 고기 및 L-아미노산)[29–31]에 의해 활성화되고, TAS1R2 및 TAS1R3 서브유닛의 이종이량체 수용체는 단맛 미각 물질(예: 자당, 과당 및 포도당과 수크랄로스와 같은 인공 감미료)에 의해 활성화된다 [30, 32–34].
- TAS1R3이 결핍된 마우스는 단맛과 우마미 맛 모두에 대한 반응이 감소했는데, 이는 이 이종이량체 복합체가 이러한 맛의 주요 매개체일 수 있지만 단맛과 우마미 맛 지각에 대한 다른 메커니즘이 존재한다는 것을 나타낸다 [35].
- 쓴맛 물질(예: 카페인, 퀴닌, 데나토늄 벤조에이트)은
⇒ 약 30개의 멤버를 가진
Type 2 taste receptor (TAS2R) 패밀리의 GPCR에 의해 감지된다 [36–38]. - 각 type II TBC는
⇒ TAS1R 또는 TAS2R 패밀리의 특정 멤버(쓴맛을 느끼는 각 세포는 4~11개의 TAS2R을 공동 발현할 수 있음)을
발현하므로
⇒ 단맛과 감칠맛, 즉 쓴맛 물질 중 하나에만 반응한다. - Type II cells는
⇒ TBC에서 합성되는 다양한 호르몬의 대부분과
그 관련 수용체(cognate receptors)를 포함한다 (Figure 2) [14,15,39–44]. - Type II TBC에서
⇒ 장쇄 지방산(long-chain fatty acids, LCFAs)을 감지하는
⇒ 유리지방산 수용체 4 (GPR120) 및
혈소판 당단백질 (platelet glycoprotein) 4 (CD36)와 같은
지방 센서(fat sensors)의 발현이 보고된 바 있다 (Figure 2) [16,17].
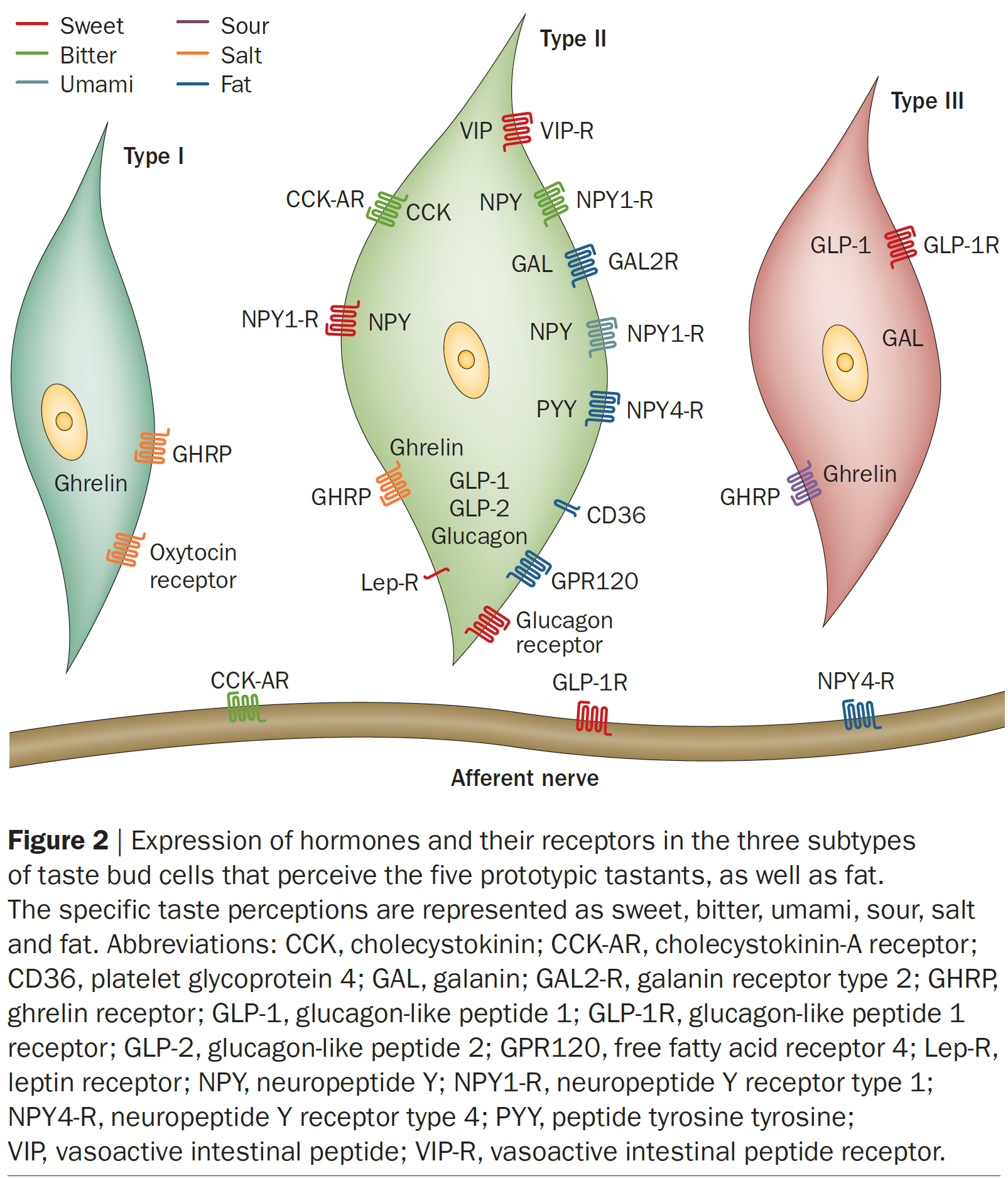
▣ Figure 2 | 다섯 가지 原型 미각 물질들과 脂肪을 감지하는 세 가지 하위 유형의 미뢰 세포들에서의
호르몬과 그 수용체의 발현.
구체적인 맛 지각은 단맛, 쓴맛, 우마미, 신맛, 짠맛, 지방으로 제시되어 있다.
약어:
CCK ⇒ cholecystokinin (콜레시스토키닌);
CCK-AR ⇒ cholecystokinin‑A receptor (콜레시스토키닌‑A 수용체);
CD36 ⇒ platelet glycoprotein 4 (혈소판 당단백질 4);
GAL ⇒ galanin (갈라닌);
GAL2‑R ⇒ galanin receptor type 2 (갈라닌 수용체 2형);
GHRP ⇒ ghrelin receptor (그렐린 수용체);
GLP-1 ⇒ glucagon-like peptide 1 (글루카곤 유사 펩타이드 1);
GLP-1R ⇒ glucagon-like peptide 1 receptor (글루카곤 유사 펩타이드 1 수용체);
GLP‑2 ⇒ glucagon-like peptide 2 (글루카곤 유사 펩타이드 2);
GPR120 ⇒ free fatty acid receptor 4 (유리 지방산 수용체 4);
Lep‑R ⇒ leptin receptor (렙틴 수용체);
NPY ⇒ neuropeptide Y (신경펩타이드 Y);
NPY1‑R ⇒ neuropeptide Y receptor type 1 (신경펩타이드 Y 수용체 1형);
NPY4‑R ⇒ neuropeptide Y receptor type 4 (신경펩타이드 Y 수용체 4형);
PYY ⇒ peptide tyrosine tyrosine (펩타이드 티로신 티로신);
VIP ⇒ vasoactive intestinal peptide (혈관활성 장내 펩타이드);
VIP‑R ⇒ vasoactive intestinal peptide receptor (혈관활성 장내 펩타이드 수용체).
Type III cells
- Presynaptic cells (type III cells)는
⇒ 감각 구심성 돌기내 신경 섬유(afferent intragemmal nerve fibres)를 가진
⇒ 통상적인 신경 시냅스를 형성하는 유일한 TBCs의 유형이다. - 뉴런(neurons)과 유사하게, 이 세포는
⇒ voltage-gated Ca2+ channels을 포함하고 있으며,
⇒ 消極(depolarized)되면
◆ 小胞性 세로토닌 (vesicular serotonin),
◆ 아세틸콜린 (acetylcholine),
◆ 노르에피네프린 (norepinephrine),
◆ γ‑아미노부티르산 (γ‑aminobutyric acid)(GABA)을 방출한다 [45]. - 이 세포들은 또한
⇒ polycystic kidney disease 2‑like 1 protein (PKD2L1, 多囊性 腎臟疾患 2형-유사 1 단백질)과
⇒ polycystic kidney disease like 3 protein (PKD1L3, 다낭성 신장질환 유사 3 단백질) 채널을 발현하는데,
⇒ 이 두 단백질은 함께 신맛(酸味)을 감지하는 데 관여한다 [46]. - 마우스에서 PKD2L1을 발현하는 type III cells가 결핍되면 산성 화학물질(예: 구연산)에 대한 반응이 완전히 사라지거나 민감도가 감소했다 [47,48].
- 높은 염도는 혐오감을 유발하며,
⇒ type II cells에서는 쓴맛을,
⇒ type III cells에서는 신맛을 감지하는 것을 활성화한다 [49].
Taste cell precursors
- TBC의 마지막 서브타입(이전에는 type IV cells라고 불림)은
⇒ 미뢰 구조의 기저부에 위치한 작고 이질적인 세포 집단으로 구성된다. - 이 세포들은
⇒ 처음에는 분화된 TBC 타입들의 유일한 전구 세포(progenitor cells, 幹細胞)로 여겨졌지만 [50],
⇒ TBC 줄기 세포 니치(stem cell niche)가
미뢰 기저부에만 위치한다는 생각은 바뀌어 더 이상은 아니다 [51–53]. - Sonic hedgehog protein (SHH, 소닉 헤지혹 단백질)이 TBCs의 분화(differentiation)를 조절한다.
- 미뢰 내 SHH-expressing cells (SHH-발현 세포)는
⇒ 미뢰 외부에 위치하며,
전사 인자(transcription factors)인 징크 핑거 단백질(zinc finger protein) GLI1과
패치 도메인 함유 1(patched 1로 알려짐)을 발현하는
SHH-expressing cells (반응 세포)에 신호를 보낸다. - 성체 마우스 혀에서는 SHH 반응 세포의 여러 영역이 미뢰를 둘러싸고 있다 [52].
- 계통 추적 실험을 통해 미뢰 내 SHH 발현 세포가 다른 세 가지 세포 유형의 직접적인 전구 세포임이 밝혀졌다 [51].
- 또한, 또 다른 연구에서는 TBCs의 전구 세포(progenitors)가 미뢰 자체 외부에 위치한다는 것을 확인했는데, 미뢰 내부에서 증식하는 세포가 매우 적거나(<10%), 전혀 없다는 것을 보여주었다 [53].
- 따라서 미뢰의 기저부에 존재하는 작은 세포는
⇒ 정지 상태의 전구 세포(quiescent precursor cells)와
미성숙 미세포(immature taste cells)의 두 가지 다른 범주로 나뉘며,
둘 다 전구 세포가 아니다. - 따라서 특정 TBC 타입을 설명하는데 type IV cell이라는 용어는 더 이상 일반적으로 사용되지 않는다.
Signalling mechanisms of taste perception
맛 지각의 신호전달 메카니즘
Type III cells가 통상적인 신경 시냅스를 갖는 유일한 미각 세포라는 점을 감안할 때 (Figure 1a), type II 및 type III cells는 간극 접합(gap junctions)을 통해 통신하여 미각 신경을 활성화시켜 뇌에 미각 물질의 특성에 관한 정보를 전달하는 것으로 이전에는 생각되었다 [54,55]. TAS1R3 프로모터의 컨트롤 하에 형광 표지된 trans-synaptic protein (트랜스 시냅스 단백질)(wheat germ agglutinin 小麥胚芽凝集素; WGA)을 발현하는 형질 전환 마우스(transgenic mice)에서, serotonin-positive cells (아마도 type III cells)에서 일부 WGA 발현이 발견되었으며, 이는 type II cells에서 단백질의 직접적 통신과 통과를 의미한다 [56].
그러나 이후 두 편의 논문에서는 type III cells에서 TAS1R3와 연결된 비형광 표지 WGA의 측면 이동(lateral transfer)을 발견하지 못했다 [57,54]. 따라서 type III cells에서만 발현되는 PKD1L3 promoter의 제어 하에 WGA를 발현하는 형질전환 마우스에서는 type II cells에 WGA가 존재하지 않았다 [55]. 이러한 데이터는 type II cells와 type III cells 간에 단백질의 직접적인 이동이 일어나지 않음을 시사한다. 더욱이, 마우스에서 type III cells를 유전자적으로 제거해도 단맛, 쓴맛 또는 감칠맛 감각이 손상되지 않았으며, 이는 이러한 세포들이 type II cells로부터의 맛 정보 전달에 필요하지 않음을 보여준다 [47].
지난 10년 동안 발표된 연구에 따르면 type II cells는 ATP 방출과 신경 섬유상의 퓨린 수용체(purinergic receptors) 활성화를 통해 미각 신경과 직접 통신하며, 따라서 두 세포 유형 간에 분자를 직접 전달할 필요가 없다 [12,24]. ATP 외에도 type II cells는 이웃 세포에 정보를 전달하기 위해 국소적으로 생성되는 호르몬을 방출한다(paracrine effect, 파라크린 효과, 주변분비 효과). 이러한 호르몬들은 호르몬을 발현하는 세포의 미각 신호 전달 메커니즘을 수정하여 자가분비(autocrine) 방식으로 맛 지각을 변경할 수도 있다 [58]. 또한 미뢰의 구심성 신경 섬유(afferent nerve fibres)에는 GLP-1 및 neuropeptide Y (NPY, 신경펩타이드 Y)와 같은 국소적으로 생성되는 호르몬에 대한 수용체가 포함되어 있어 [14,59], 특정 미각 물질에 노출된 후 하위 세포 신호 전달의 '온 스위치' 역할을 하는 미각 신호의 강도를 조절할 수 있다 (Figure 2).
TAS1Rs (umami and sweet), TAS2Rs (bitter) 그리고 지방 수용체(fat receptors)는 TBC의 서로 다른 서브타입에 속하지만, 다운스트림 신호 전달 경로를 공유하며, 궁극적으로 ATP와 호르몬의 방출을 유도한다. 구체적으로, 미각 물질 (단맛, 쓴맛, 감칠맛 또는 LCFAs)이 특정 테이스트 수용체 또는 센서에 결합하면 PLCβ2가 활성화되고 IP3와 같은 2차 전달물질이 생성된다. IP3는 TRPM5 (transient receptor potential cation channel subfamily M member 5, 일시적 수용체 전위 양이온 채널 서브패밀리 M 멤버 5; Trpm5에 의해 인코딩됨) 채널을 여는 세포 내 Ca2+ 방출을 유발하여 세포 소극을 초래한다 [60–62].
TBC에서 생성된 활동 전위(action potentials)는 전압 개폐형(voltage-gated) CALMH1 (calcium homeostasis modulator 1, 칼슘 항상성 조절자 1) 채널을 통해 [63,64], non-vesicular ATP (비소포 ATP)의 방출로 이어지며, 이 채널은 감각 신경 섬유와 type II 및 type III cells의 퓨린 수용체를 활성화한다 [12]. Type II cells에서 퓨린 수용체의 활성화는 ATP의 증가된 방출을 강화하는 반면, type III cells에서는 전압 개폐성 Ca2+ 채널이 활성화되어 고전적인 신경전달물질의 방출을 유발한다 [12,65]. 마우스에서 Trpm5의 녹아웃은 단맛, 쓴맛 및 우마미 구별을 없앴고 [66], Trpm5는 또한 TBCs의 LCFA-mediated depolarization (LCFA-매개 소극)과 지방이 없는 미각 물질보다 지방에 대한 마우스의 선호도에 필요한 것으로 밝혀졌다 [61,67,68]. 마우스에서 CALHM1의 녹아웃은 단맛, 우마미 및 쓴맛에 대한 지각을 심각하게 손상시켰을 뿐만 아니라 미각 물질에 대한 반응으로 type I TBC에서 ATP 방출이 크게 감소했다 [64].
이러한 관찰 결과를 종합해 볼 때, type II cells로부터 뇌로 미각 정보가 전달되는 주요 경로는 ATP 방출을 통해 이루어지며, ATP는 미각 신경과 직접 상호작용하여 고차 뉴런(higher order neurons)으로 정보를 전달한다. 중요한 점은 type II cells로부터 방출된 ATP가 type I cells의 membrane-bound (막-결합) ATPase에 의해 분해되어 ADP를 생성하고, 이는 구심성 신경섬유의 퓨린성 수용체 탈감각(desensitization)을 방지한다는 것이다 [69]. 비특이적 엑토펩티다아제(Non-specific ectopeptidases)는 일부 ADP 분자를 아데노신(adenosine)으로 분해하여 아데노신 수용체 A2B에 작용할 수 있도록 하는데, 이 수용체는 TAS1R을 발현하는 세포 일부에 존재하며, 단맛을 내는 리간드(ligands)에 의한 TAS1R의 활성화에 반응하여 ATP 방출을 증가시킨다 (아마도 호르몬 방출도 증가시킬 것임) [70].
신맛(Sour taste)은 양성자가 type III cells에 유입되어 세포의 산성화를 유발할 때 감지된다. 이러한 양성자 유입 (proton influx)은 resting K+ 채널의 폐쇄, 막 소극(membrane depolarization) 및 고전적 신경전달물질의 방출을 초래한다 [71].
Hormones expressed in taste buds
미뢰에서 발현되는 호르몬들
CCK and Y‑family peptides
CCK와 NPY는 쥐 TBCs에서 처음으로 기술된 호르몬이었다 [41,72]. CCK는 콜레시스토키닌(cholecystokinin) 수용체 type A (CCK‑AR)을 통해 자가분비(autocrine) 방식으로 작용하고, 지연 정류기(delayed rectifier)와 내향 정류기(inward rectifier) K+ 채널 모두의 활성화를 억제하며, TAS2R 자극 시 세포 내 Ca2+를 최대 30%까지 증가시킨다 [41]. K+ 채널이 TBC의 막 재분극(membrane repolarization)에 필요하다는 점을 감안할 때 이러한 세포는 더 오랜 시간 동안 탈분극 상태(depolarized state)를 유지하며 이로 인해 오래 계속되는 쓴맛 신호전달(prolonged bitter signalling)을 초래한다 [58]. 또한 CCK‑AR이 결핍된 쥐는 인공 감미료를 포함한 달콤한 물질에 대한 핥기 반응이 증가하는데, 이는 이 수용체 부재가 맛 지각을 변화시킨다는 것을 보여준다 [73].
TBCs에서 NPY의 발현은 CCK의 발현과 거의 완전히 겹친다; 마찬가지로 NPY는 막 탈분극 시 TBC에서도 방출된다 [58].
Y 펩타이드 패밀리는 췌장 폴리펩타이드(pancreatic polypeptide), NPY, 그리고 peptide tyrosine tyrosine (PYY)으로 구성되며, 이는 NPY 수용체(NPY1‑R, R, R, NPY5‑ R and R)로 알려진 GPCR 패밀리를 통해 에너지 대사를 조절하는 데 관여한다 [74]. NPY4‑R은 미각 신경 섬유에, NPY1‑R은 TAS1Rs을 발현하는 type II TBC와 미세융모 돌기 (microvilli projections)에 발현된다 [58,59]. TBC에서 NPY는 CCK에 길항적으로 작용하여 inwardly rectifying K+ currents (내향 정류 K+ 흐름)를 증가시켜 단맛과 감칠맛에 반응하는 신호 전달을 하향 조절(downregulation)한다 [72,75]. 쓴맛에 대한 지각을 증폭시키는 이러한 계층적 신호 전달 체계는 단맛 자극과 그에 수반되는 쾌락 감각이 동시에 제시되어도 쓴맛 독소가 가려지지 않도록 한다. 야생에서 단맛과 쓴맛을 나란히 제시했을 때 영장류가 쓴맛(유독하거나 썩은 식물)과 단맛(익고 일반적으로 독성이 없는 식물)을 구별할 수 있도록 하는 능력은 진화적으로 생존에 매우 중요하다 [76]. 이 과정은 쓴맛이 가장 중요하다는 것을 보장하는 여러 적응 메커니즘 중 하나일 뿐이다.[77]
NPY1‑R 및 NPY4‑R을 활성화하는 PYY는 L‑cells에서 분비되며 미뢰에서도 생성된다 [43]. 설치류(Rodents)는 두 가지 형태의 단백질을 발현한다: 모든 NPY 수용체에 대해 비교적 비선택적인 PYY(1–36)과, 그리고 DPP4 (dipeptidyl peptidase 4) 효소 활동의 산물인 PYY(3‑36)는 순환계에서 발견되는 가장 풍부한 단백질 형태이며, NPY2‑R을 우선적으로 활성화한다 [78]. TBC는 DPP4를 발현하지 않는다 [14]. 따라서 PYY(1–36)은 미뢰에 존재하는 가장 풍부한 단백질 형태일 가능성이 높다. PYY가 결핍된 마우스는 지방 맛에 대한 선호도가 상당히 감소한 것으로 나타났다 [43,59]. 또한, CD36 (포화 및 불포화 LCFA를 결합(bind)하는 스캐빈저 수용체 계열(scavenger receptor family)에 속하는 지질 센서(lipid sensor))이 결핍된 마우스[79,80]는 LCFA가 풍부한 용액과 고형 음식에 대한 맛 선호도가 전혀 나타나지 않았다. 이는 CD36 기능이 LCFAs의 구강 구별(oral discrimination)에 절대적으로 필요하다는 것을 나타낸다 [81]. PYY는 CD36 발현 및/또는 기능 조절에 관여할 수 있지만 이러한 가능성은 아직 연구되지 않았다.
Glucagon, GLP‑1 and GLP-2
Glucagon, GLP‑1, 그리고 glucagon-like peptide 2 (GLP-2)는 GCG에 의해 인코딩된 single proglucagon peptide에서 생성된 분열 산물(cleavage products)이다 [82]. GLP‑1 및 GLP‑2는 장내분비(enteroendocrine) L‑cells에서 발견되는 주요 단백질 산물이며, 글루카곤은 췌장(pancreas)의 랑게르한스 섬(islets of Langerhans)에 있는 α‑세포에서 생성되는 주요 펩타이드이지만 GLP‑1 및 GLP‑2는 이 세포에서도 생성된다 [83]. Glucagon-like hormones은 TAS1R을 발현하는 type II cells와 일부 type III cells에 존재하며 이들의 동종 수용체(cognate receptors)는 인접한 구심 섬유에서 발현된다 (Figure 2).[14] GLP‑2는 장의 강력한 영양 요소(trophic factor)이다 [84]. 그러나 이 호르몬이 TBC에서 하는 역할은 알려져 있지 않다. 이에 비해 랑게르한스 섬에서 인슐린 분비를 촉진하는 GLP‑1은 단맛에 기여한다 [82].
GLP‑1 수용체(1R)가 결핍된 마우스는 칼로리-함유 감미료(sucrose)와 인공 감미료(sucralose)에 대한 미각 반응이 극적으로 감소하고, 감미료에 대한 민감도가 증가한다 [14,85]. Sucrose, 인공 감미료 및 umami 자극은 마우스의 有郭乳頭(circumvallate papillae)에 있는 TBCs로부터의 GLP‑1과 NPY의 분비를 유발한다. 이는 TAS1R3 결핍 마우스에서는 나타나지 않는 효과이다 [86]. 이 발견은 하류 신호 전달, 세포 소극 및 GLP‑1R로의 신호 전달이 주어진 미각 물질의 완전한 지각을 위해 감각 구심성 신경에 의존함을 보여준다. 신맛과 높은 솔트 농도는 TBCs로부터의 NPY 분비를 증가시키지만, GLP‑1 분비는 증가시키지 않는다.[87] 인간과 마우스들을 대상으로 한 연구에 따르면, LCFAs는 GLP‑1R 의존적 방식으로 sucrose에 대한 선호도를 강화하는 것 외에도 TBCs에서 GLP‑1 분비를 증가시키는 것으로 나타났으며, 이는 아마도 GPR120과의 상호작용에 의해서일 것이다.[88,89] 활성 GLP‑1을 빠르게 분해하는 효소인 DPP4가 미뢰에 존재하지 않는다는 점을 감안할 때, GLP‑1의 높은 국소적 농도로 인해 GLP‑1R 신호가 증가할 가능성이 있다 [14]. TBCs에서의 GLP‑1과 NPY의 발현은 단맛, 감칠맛, 쓴맛 및 신맛(혐오)을 구별하기에 충분하다 [87]. 글루카곤(Glucagon)과 그 수용체는 type II cells의 하위 집단에 존재하며(Figure 2) [42], TBCs에서 글루카곤 신호의 약리학적 또는 유전적 파괴는 단맛 신호만 감소시켰다 [42]. 특정 수용체를 통한 측분비(Paracrine) 글루카곤 신호전달은 국소 회로(local circuit)에서 단맛 신호전달을 증강시킨다. 그러나 단맛 미각 물질이 GLP-1 release에 미치는 영향과는 달리, 단맛 미각 물질에 반응하여 글루카곤 분비가 감소한다. 이는 글루카곤이 GLP-1과 GLP-2를 발현하는 것들 보다는 type 2 TBCs의 다른 집단에서 발현됨을 시사한다.
VIP peptides
VIP (Vasoactive intestinal polypeptide, 혈관활성 장 펩타이드)는 VIP 수용체인 VIP‑R-1 및 VIP‑R-2 외에도 TAS1R3 및 TAS2R을 발현하는 세포들의 하위집단(subpopulation)에서 합성된다 [40].
VIP가 결핍된 마우스는 단맛과 쓴맛 물질에 대한 지각이 향상된다. 따라서 GLP‑1 신호전달 경로의 활성화는 VIP 신호전달 경로와 비교하여 단맛 지각에 반대되는 효과를 나타낸다. 흥미롭게도, GLP‑1을 발현하는 세포의 수와 이 호르몬의 전반적인 발현 수준은 VIP가 결핍된 마우스의 TBCs에서 상당히 증가했는데, 이는 GLP‑1 또는 VIP를 발현하는 세포의 수에 내재적인 균형이 존재함을 시사한다 [40].
Ghrelin
식욕 조절의 맥락에서 널리 연구된 그렐린(ghrelin)은 위(stomach)의 X/A-like cells에서 분비되며 그 수용체인 GHRP와 함께 4가지 TBC 서브타입들 모두에서 발현되는 것으로 보고되었다 [15]. 그러나 그렐린을 아실화하고(acylates) 활성화하는 효소인 ghrelin‑O-acyltransferase (GOAT, 그렐린-O-아실트랜스퍼라제)의 발현은 그렐린을 발현하는 세포의 하위 집합으로 제한되는 것으로 보인다 [15,90,91]. GHRP가 결핍된 마우스는 짠맛과 신맛에 대한 지각이 상당히 감소한다; 그러나 아실화된 그렐린 외에도 GHRP와 리간드 이외의 수용체가 TBCs에서 이러한 반응을 주도할 수 있다 [92,93]. GOAT 또는 ghrelin이 결핍된 마우스는 맛 지각과 관련하여 약간 다른 표현형을 보였다. PYY-결핍과 유사하게, GOAT 또는 그렐린 결핍은 LCFAs에 대한 반응성을 감소시켰다 (이는 이 경로가 PYY 경로와 상승적으로 작용할 가능성을 시사한다). 그러나 그렐린 결핍 마우스에서는 짠맛에 대한 지각이 감소했지만, GOAT 결핍 마우스에서는 짠맛에 대한 지각이 향상되었다 [15].
Oxytocin
TBCs와 TBCs를 둘러싼 신경 섬유들에는 옥시토신(oxytocin) 발현이 없는 것으로 보이지만, 옥시토신 수용체는 type I (low-salt sensing) TBCs에 존재한다 (Figure 2). Type I TBCs를 옥시토신에 노출시키면 Ca2+ 이동이 발생하는데, 이는 옥시토신 수용체 길항제에 의해 차단된다 [94]. 이러한 조직학적(histological) 및 세포학적 발견들은 옥시토신이 염분 식욕(salt appetite)과 나트륨 尿(natriuresis)를 조절하는 호르몬으로서 이미 알려져 있는 역할을 뒷받침한다.
Galanin
주로 중추 신경계와 腸에서 발견되는 갈라닌(galanin)[96]은 음식 섭취, 뿐만 아니라 장 운동성(gut motility) 및 호르몬 분비(hormone secretion)를 조절하는 데 관여한다 [97]. 이 호르몬은 type II 및 type III TBCs에서 발현된다; 갈라닌 수용체 type 2 (GAL2‑R)을 코딩하는 mRNA 전사본(transcripts)이 미뢰에서 발견되었다 (Figure 2) [98]. 갈라닌을 과발현한 마우스는 지방이 풍부한 식단의 소비가 55% 증가했지만 단맛이나 쓴맛에 대한 선호도는 증가하지 않았다 [99]. 반대로 갈라닌이 결핍된 마우스는 야생형 마우스에 비해 지방이 많은 식단에 대한 선호도가 감소했다.[100] 이러한 결과는 type II TBC에서 지질 센서 신호전달 장치(CD36 및/또는 GPR120을 통해)를 조절하여 지방 소비 조절에 갈라닌이 역할을 한다는 것을 강력하게 뒷받침한다 [88,101].
Leptin
Leptin (렙틴)은 에너지 소비, 체중, 체지방량, 그리고 섭식 행동을 조절하는 데 관여하는 핵심 호르몬이다. 순환 렙틴 (circulating leptin) 수치는 전신 비만도(body adiposity)와 높은 상관관계를 보인다 [44]. 렙틴 수용체는 type II cells에서 발현되며(Figure 2), 실온(24°C)에서 활성화되면 외부로 K+ currents가 흐르게 되어 TBCs가 단맛 자극에 대해 과분극된다 [102]. 그러나 렙틴을 투여한 후 35°C에서 마우스의 혀에 단맛 자극을 가했을 때, 혀 앞쪽에 있는 TBCs로부터 미각 정보를 전달하는 제7 뇌신경(cranial nerve)(안면신경)의 鼓索神經 (chorda tympani) 가지를 통한 신경 반응의 크기가 증가했다 [103]. 연구의 실험 조건은 달랐지만 TRPM5 채널은 온도가 증가함에 따라 활성화가 증가하는 것으로 알려져 있다 [104]. 따라서 이러한 결과는 온도가 증가함에 따라 감각 신경을 통한 활동이 증가하고 ATP 방출이 증가하는 TRPM5 채널의 활동을 증가시키는 렙틴의 역할을 뒷받침한다.
※ 고실끈신경, 고삭신경(鼓索神經). 안면신경(중간신경)으로부터 출발하여 下顎, 舌下腺 그리고 舌腺에 분포하고 혀 앞부분의 2/3를 지배하고 있는 신경. 副交感神經과 특수 감각신경이다.
렙틴이 단맛 지각을 조절한다는 가설은 정상 마우스와 유전적으로 렙틴이 결핍된 마우스에게 렙틴을 투여했을 때, sucrose와 saccharin에 대한 행동 검사 반응이 상당히 감소했다는 관찰 결과에 의해 뒷받침된다. 그러나 렙틴 수용체가 결핍된 생쥐에게 렙틴을 투여한 후에는 행동 미각 변화가 나타나지 않았다 [44]. 결과적으로, 렙틴에 민감한 상태에서는 렙틴이 단맛이 함유된 음식의 과식을 막는 역할을 할 수 있다. 그러나 비만과 같은 렙틴 저항성 상태에서는 이러한 음식 섭취 조절 기전이 약화될 수 있다. VIP가 결핍된 생쥐는 단맛 지각이 증가하고 GLP-1을 발현하는 TBCs의 수가 증가하지만 순환 렙틴 수치는 감소한다는 점을 고려할 때, 이러한 데이터는 미뢰에서 VIP, GLP-1, 그리고 leptin 활성 사이에 항상성 및 기능적 상호작용이 존재함을 뒷받침한다 [40].
TBCs have similarities to islets of Langerhans
미뢰와 랑게르한스섬(췌장의 내분비 세포)은 표현형적으로 많은 유사성을 공유한다. 구조적으로 두 세포 모두 상피 조직에 내장된 내분비 기관이며, 주변 실질조직(parenchyma)과는 다른 특수한 기능을 가지고 있으며, 자체적인 혈액 공급을 받는다 [105, 106]. 또한, 랑게르한스섬과 유사하게, 일부 TBCs는 연관된 특정 호르몬에 대한 동족 수용체를 공동 발현한다 [107]. 이와 대조적으로, 랑게르한스섬과 미뢰는 소화 과정의 여러 단계에서 영양소의 섭취와 처리를 조절한다. 미뢰는 음식의 다양한 성분으로부터 감지된 정보를 통합하는 기관으로, 음식 성분에 대한 흡수 전 반응(preabsorptive responses)에 기여한다 [108]. 반면, 랑게르한스섬 호르몬 분비는 주로 음식 섭취 후 흡수된 산물에 의해 조절되어 에너지 이용과 포도당 대사를 조절하는 기능을 한다 [109].
The gut as a chemosensory organ
화학감각기관으로서의 장
腸은 세포 수와 호르몬 다양성 측면에서 신체에서 가장 큰 호르몬 생성 기관이지만, 장내분비 세포는 전체 장 상피 (intestinal epithelium)의 1%에 불과하다 [110]. 분자 센서 역할을 하는 위장관 상피 세포 (Gastrointestinal epithelial cells)는 음식 섭취 및 소화와 관련된 여러 과정에 관여한다 [111]. 흥미롭게도 장에서 확인된 많은 호르몬(Figure 3)이 TBCs에서도 발현된다.

▣ Figure 3 | 腸을 따라 분포하는 특정 호르몬들. 腸에서 발현되고 미뢰 내 미뢰 세포에도 존재하는 호르몬들.
CCK와 그렐린은
⇒ 上部 胃腸管 (upper gastrointestinal tract)에서 발견된다.
⇒ CCK는 I-세포에서 분비되고
⇒ 그렐린은 X/A-유사 세포에서 분비된다.
中部 胃腸管(middle gastrointestinal tract)에서 K-세포는 ⇒ GIP를 분비한다.
下部 胃腸管(lower gastrointestinal tract)에서 L-세포는 ⇒ PYY, GLP-1 및 GLP-2를 분비한다.
약어:
CCK ⇒ cholecystokinin (콜레시스토키닌);
GIP ⇒ gastric inhibitory polypeptide (위 억제 폴리펩티드);
GLP-1 ⇒ glucagon-like peptide 1 (글루카곤 유사 펩티드 1);
GLP-2 ⇒ glucagon-like peptide 2 ( 글루카곤 유사 펩티드 2);
PYY ⇒ peptide tyrosine tyrosine (펩티드 티로신 티로신).
Sweet taste receptors
단맛 수용체
腸의 영양소 감지(nutrient sensing)가 소화 뿐만 아니라 음식 섭취 조절에도 중요한 바, 연구 노력들은 taste-gut-brain 축을 이해하고 이 축이 음식 섭취를 조절하는 기능을 하는 방식에 초점을 맞춰 왔다. 영양소에 의해 직접 활성화되는 TAS1Rs는 胃의 brush cells, K-cells, L-cells, K/L enteroendocrine cells (장내분비세포), X/A-like cells에서 발견되었다 [112]. Glucose (포도당)이나 sucralose에 노출되면 마우스 소장 세포(small intestine)에서 GLP-1과 GLP-2 분비가 증가하는데, 이는 단맛 억제제인 구르마린(gurmarin)으로 상쇄할 수 있다 [113]. 장내 TAS1R은 장 호르몬 분비와 관련이 있으며, guanine nucleotide-binding protein G(t) subunit α‑3 (type II TBCs에서 발현되는 특정 GTP 결합 α‑ 서브유닛; 일반적으로 α‑gustducin으로 알려짐)이 결핍된 마우스에서는 포도당 매개 GLP‑1 방출이 감소한다 [114]. 이는 장내 TAS1R이 포도당 센서일 수 있음을 시사한다 (Figure 4). TAS1R2/TAS1R3 heterodimers (이종이량체)와 α‑gustducin은 GLP‑1과 gastric inhibitory polypeptide (위 억제 폴리펩타이드)를 모두 발현하는 K/L cells와, 1 및 2를 발현하는 L-cells에 존재한다 [115,116].
포도당-매개 GLP‑1 방출은 여러 기전에 의해 조절된다: 대표적인 두 가지 기전은, 췌장의 β 세포와 같은 포도당 감지 기전과 sodium/glucose transporters (SGLT)에 의해 조정되는 sodium-coupled glucose uptake (나트륨-결합 포도당 흡수)이다 [117].
SGLT‑1 단백질은 식이 당을 장 내강(intestinal lumen)에서 장세포(enterocytes)로 운반하는 주요 경로이다. TAS1R은 내강 당(luminal sugars)과 감미료에 반응하여 SGLT‑1의 상향 조절에 상당한 역할을 한다. TAS1R3 또는 α‑gustducin이 결핍된 쥐는 탄수화물이 함유된 음식이 소장으로 유입되는 것에 반응하여 장에서 1 단백질의 발현을 증가시킬 수 없다. 이러한 동물에서는 설탕 섭취 후 나트륨-포도당 의존적 수송도 손상된다 [115].

▣ Figure 4 | 味覺器官과 器官系의 關係.
미각 유두(taste papillae)로부터의 미각 정보를 전달하는 신경 섬유는
⇒ 腦幹(brainstem)의 NTS (Nucleus tractus solitarius, 孤束核, 고립로핵)에서 모인다;
맛 지각 신호는
⇒ PBN(설치류에서만 나타남)으로 전달되어 視床(thalamus)을 거쳐
⇒ 섬엽(insula)의 일차 미각 피질(primary gustatory cortex)에서 끝난다.
⇒ 腦橋(pons)를 지나면 섬유의 3분의 1이 視床에서 교차하여 섬엽(insula)에서 양쪽으로 미각을 표상한다.
NTS로부터의 국소 돌기(Local projections)는
⇒ 타액선(salivary glands)의 타액 분비(salivation)와
폰 에브너선 (Von Ebner’s glands)의 장액 분비(serous secretions)를 조정한다.
α‑gustducin은
⇒ 膵管(pancreatic ducts)과 창자(intestine)의 솔세포(brush cells)에서 발현되지만,
⇒ 이 세포에서의 기능은 아직 밝혀지지 않았다.
α‑gustducin과 TAS1R은
⇒ 랑게르한스섬의 β세포에서도 발현되며,
⇒ 기저 인슐린 분비(basal insulin secretion) 조절에 관여할 가능성이 있다.
또한, 膵島(islets)의 TAS1R은
⇒ 果糖(fructose)에 의해 활성화되어
⇒ 포도당-매개 인슐린 분비(glucose-mediated insulin secretion)가 증가한다.
腸內分泌 細胞(enteroendocrine cells)에서
⇒ 단맛 수용체 (TAS1R2/TAS1R3 또는 추정 TAS1R3/TAS1R3 이합체),
⇒ 우마미 수용체 (TAS1R1/TAS1R3),
⇒ 쓴맛 수용체 (TAS2R) 및 장쇄 지방산에 대한 수용체 (GPR120, GPR40 및 CD36)가 발현되고,
⇒ LP‑1, CCK 및 그렐린의 분비에 관여한다.
포도당과 지방은 장세포(enterocytes)로 흡수된다:
⇒ 포도당 흡수는 나트륨-의존성 SGLT1 채널을 통해 이루어지고,
⇒ 지방 분자는 내경 표면(luminal surface)을 통해 자유롭게 확산된다.
약어:
CCK ⇒ cholecystokinin (콜레시스토키닌);
CD36 ⇒ platelet glycoprotein 4 (혈소판 당단백질 4);
GLP‑1 ⇒ glucagon-like peptide 1 (글루카곤 유사 펩타이드 1);
GPR40 ⇒ free fatty acid receptor 1 (유리 지방산 수용체 1);
GPR120 ⇒ free fatty acid receptor 4 (유리 지방산 수용체 4);
NTS ⇒ nucleus tractus solitarius (孤立核);
PBN ⇒ parabrachial nucleus (팥곁핵, 상완핵, 부완핵);
SGLT1 ⇒ sodium/glucose cotransporter 1 (나트륨/포도당 공동수송체 1);
TAS1R1 ⇒ taste receptor type 1 member 1 (미각 수용체 1형 구성원 1);
TAS1R2 ⇒ taste receptor type 1 member 2 (미각 수용체 1형 구성원 2);
TAS1R3 ⇒ taste receptor type 1 member 3 (미각 수용체 1형 구성원 3);
TAS2R ⇒ taste receptor type 2 (미각 수용체 2형).
Bitter taste receptors
쓴맛 수용체
TAS2R 패밀리의 여러 멤버들은 포유류 장(mammalian intestine)과 마우스 장에서 유래한 enteroendocrine cell line (장내분비 세포라인) STC‑1 (CCK와 proglucagon 산물들을 합성하는 능력으로 인해 장내분비 세포의 분비 기전을 연구하는 데 사용되는 모델 시스템)에서 발현된다 [111,118]. 마우스에서 쓴맛 수용체는 結腸 (colon)과 胃(stomach)의 일부 杯狀細胞(goblet cells)에 국한되어 있다. 그러나 結腸에서의 기능이 있는지 여부는 아직 밝혀지지 않았다 [119].
야생형 마우스에 原型(prototypic) TAS2R-작용제 혼합물(經口 胃管營養, oral gavage)을 투여한 결과, 위관영양 40분 후 최고 농도에 도달하는 아실화 그렐린(acylated ghrelin)의 혈장 레벨이 증가했다. 아실化 그렐린의 분비는 α-gustducin (X/A-like cells에서 발현)에 의해 조절되는 것으로 보이는데, α-gustducin 결핍 마우스는 쓴맛이 나는 혼합물에 대한 반응이 둔화되었기 때문이다. 또한, 야생형 마우스에서 쓴맛이 나는 혼합물 위관영양으로 인한 아실화 그렐린 분비량의 증가는 음식 섭취량의 급격한 증가로 이어졌고, 이후 4시간 동안 음식 섭취량이 감소했으며, 이는 위 배출 지연(delay in gastric emptying)과 관련이 있었다. 이러한 효과는 유사하게 위관영양을 투여한 α-gustducin 결핍 마우스에서는 관찰되지 않았다 [120]. 이러한 위 배출 지연은 잠재적으로 독성이 있는 화학물질이 소장으로 유입되어 섭취되는 것을 방지할 수 있다. 또한, 위장 내 독소 잔류는 구토 가능성을 증가시킬 가능성이 높다. 흥미롭게도 설치류에게 데나토늄 벤조에이트(denatonium benzoate)를 위내 주입한 결과, 다른 쓴맛 화합물의 섭취를 막는 맛 혐오감이 나타났다 [121]. 따라서 장내 쓴맛 수용체는 후각 및/또는 미각 보호 기전을 우회하는 독성 화합물을 감지하기 위해 존재할 수 있다. 이러한 생각은 TAS2 수용체 작용제가 포만감을 연장하고, 식사 간격을 늘리고, 특정 병용 식품에 대한 혐오감을 유발할 수 있으므로 비만 치료에 유용할 가능성을 제기한다.
Fat-sensing receptors
지방 감지 수용체
LCFA는 친유성 특성(lipophilic nature)으로 인해, 이전에는 세포막을 자유롭게 통과하는 것으로 여겨졌다. 그러나 장에 지방 수송체가 존재한다는 새로운 증거는 LCFA 흡수가 활성적인 세포 과정임을 시사한다. 지방 흡수 초기 단계에 관여하는 후보 중 하나는 type II TBCs에서 발현되고 장(주로 십이지장 (duodenum), 空腸(jejunum), 回腸(ileum))을 따라 차등적으로 발현되는 scavenger receptor CD36이다 (위와 대장에서는 덜 풍부함) [118].
LCFA를 胃腸管(gastrointestinal tract)의 해당 부위에 직접 주입한 후, 마우스 소장(small intestine)에서 CD36을 암호화하는 mRNA 전사체 레벨이 상당히 감소했다. 이는 지방산 전이의 포화 메커니즘 (saturable mechanism of fatty acid translocation)이 존재함을 시사하며[122], 이는 섭취 상태에서 관찰되는 미뢰의 CD36 수치 감소와 일치한다 [123].
PYY가 LCFA에 반응하여 L-cells에서 분비된다는 점을 고려할 때, 이 펩타이드 호르몬이 TBCs에서 발생하는 것으로 알려진 것과 유사한 방식으로 장에서 CD36 발현을 조절하는 것으로 추정된다. CD36이 결핍된 마우스는 유사하게 처리된 야생형 마우스에 비해 위내 LCFA 투여 후 세크레틴(secretin)과 CCK의 분비가 각각 60%와 50% 감소하는 것으로 나타났다[124]. 이는 CD36 신호전달의 활성화가 LCFA-매개 호르몬 분비를 조절함을 시사한다 (Figure 4).
장 호르몬의 LCFA-매개 분비는 지방산-특이적 GPCR의 큰 계열을 통해 발생하며 [125, 126], 이 중 일부는 種 및 조직-특이적으로 발현된다. 예를 들어, free fatty acid receptor 1 (GPR40)은 마우스[127]와 인간 췌장 β 세포[128, 129], 그리고 마우스의 미뢰[130, 131]에서 우선적으로 발현되지만, 인간 혀에서는 발현되지 않는다 [132]. 이와 대조적으로, GPR120은 type II TBCs에서 발현된다 [16, 17, 123]. 이 패밀리의 수용체들은 영양소 센서 역할을 하며 에너지 항상성 유지에 중요하다 [133].
지방산-특이적 GPCR은 장 내분비세포(intestinal enteroendocrine cells)(Figure 4)에서 발현되며, 그 기능과 작용기전은 현재 진행 중인 연구의 초점이다. 10년 전, 리놀레산(linoleic acid) (α–LA)은 적어도 부분적으로는 GPR120 신호전달을 통해 1 cells와 마우스들 모두에서 GLP‑1 분비를 자극하는 것으로 나타났다 [89]. 이 발견은 TBCs에서의 LCFAs-mediated GLP‑1 secretion (LCFA-매개 GLP‑1 분비)에서 GPR120 신호전달의 역할과 일치한다 [88]. 이후 마우스를 대상으로 한 다른 연구에서도 GLP‑1 분비가 GPR120 신호전달과 무관한 것으로 보이며, GPR40이 GLP‑1 분비를 유도하는 주요 LCFA 수용체일 가능성이 있음을 시사하는 증거가 제시되었다 [134]. 보고된 결과 간의 불일치는 STC‑1 세포와 마우스에서 다양한 지방산 GPCR의 발현 차이 때문일 수 있다.
CD36 외에도 GPR120과 GPR40이 腸에서 LCFA-mediated secretion of CCK에 관여하는 것으로 보고되었다 [135,136]; 그러나 TBCs에서 CCK 분비에 관여하는 정도는 불분명하다. SCT‑1 세포에서 수행된 한 연구에서는, GPR120이 LCFA를 매개로 한 CCK 분비의 주요 매개체임을 보여주었다 [135]. 그러나 GPR40이 결핍된 마우스에서 분리한 CCK-eGFP-positive I‑ cells를 사용한 연구에서는 야생형 동물에서 분리한 동등 세포와 비교했을 때 α‑LA를 매개로 한 CCK 분비가 50% 감소한 것으로 나타났다 [136].
Endocannabinoid signalling
내인성 칸나비노이드 신로전달
Anandamide (a fatty-acid ethanolamine)와 2 arachidonoyl-sn-glycerol (2-AG)은 GPCR cannabinoid receptor 1(CB1)과 cannabinoid receptor 2 (CB2)의 내생적 리간드(endogenous ligands)이다 [137].
CB1은 type II TBCs에서 발현되며, 단맛에 대한 지각를 향상시키는 기능을 한다 [138]. central endocannabinoid system, 특히 CB1-매개 신호전달은 음식 섭취를 조절하지만, peripheral endocannabinoid mechanisms도 관여할 수 있다. 변연계 전뇌(limbic forebrain)와 視床下部(hypothalamus)의 아난다마이드(anandamide) 수치는 금식 후 증가한다 [139]. Anandamide (및 기타 CB1 작용제)를 말초에 투여한 결과, 부분적으로 포만감을 느낀 마우스의 음식 섭취가 상당히 증가했다 [140]. 놀랍게도, 5일 동안 모의 섭식(sham-feeding)한 동물에게 경구 지방(oral fat)과 완전 액상 사료(complete liquid meal)를 투여했을 때, 상부 위장관의 아난다마이드와 2-AG 수치가 증가했지만, 수크로오스와 단백질 투여는 그렇지 않았다. 迷走神經(vagus nerve)이 절단되었을 때 장 조직 내 엔도칸나비노이드(endocannabinoid) 축적이 사라졌는데, 이는 장에서 엔도칸나비노이드가 이동하려면 미주신경 자극이 필요함을 시사한다 [141,142]. 또한, 일부 추정 fat-sensing orphan GPCR은 소장에서 풍부하게 발현된다. G‑protein coupled receptor [55]와 G‑protein coupled receptor 119는 엔도칸나비노이드 수용체로 간주되며(CB1 및 CB2와 상동성이 거의 없음에도 불구하고), 장에서 엔도칸나비노이드가 이동되는 데 중요한 역할을 할 수 있다. 그러나 이들이 음식 섭취를 조절하는 기능을 하는지는 아직 밝혀지지 않았다 [143].
Role of the gut microbiota in food sensing
음식 감지에 있어서의 장 미생물군의 역할
비만(Obesity)과 대사 장애(metabolic disorders)는 장내 미생물군(gut microbiota)의 구성 및 생태 변화와 관련이 있다 [144]. 장내 미생물군의 구성은 개체마다 매우 다양하지만 건강한 개체에서는 안정적이다. 장내 미생물군이 없는 마우스(germ-free mice 무균 마우스라고 함)는 칼로리를 더 많이 섭취함에도 불구하고 정상 마우스보다 상당히 마른 편이다 [145]. 이는 미생물군이 숙주의 에너지 항상성(energy homeostasis) 조절에 기여한다는 가설을 뒷받침한다. 무균 마우스에서 혀 유두의 TBCs 형태 또는 호르몬 함량에 변화가 있는지 여부는 아직 조사되지 않았다. 그러나 TAS1R, α‑gustducin 및 1을 암호화하는 유전자의 발현 분석 결과, 무균 마우스의 장에서 대조군에 비해 증가한 반면, 혀 조직에서는 TAS1R과 α‑gustducin을 암호화하는 유전자의 발현에 차이가 관찰되지 않았다 [146]. 또한, 무균 마우스는 단맛에 대한 선호도는 변함이 없었지만, 대조군 마우스보다 고농도의 자당 용액을 더 많이 섭취했는데 [145], 이는 맛-센싱 분자와 포도당 수송체를 인코딩하는 유전자의 발현이 장내에서 상향 조절되는 것과 관련이 있다. 이러한 상향 조절은 마른 마우스에서 순환혈당(circulating blood glucose) 수치 감소(에너지 부족을 반영)에 대한 항상성 적응일 수도 있다. 따라서 무균 마우스에서 급식 및 공복 상태에서 혈당 수치를 지속적으로 모니터링하는 것은 이 모델에서 혈당 조절에 대한 이해를 높이는 데 도움이 될 수 있다.
장내 미생물에 의한 다당류 발효를 통해 생성된 단쇄 지방산은 장 상피에 위치한 GPCR의 리간드이며, 에너지 균형 유지 및 대사 장애 발생과 관련이 있는 것으로 보고되었다. 지방 인식에 있어 미생물군의 역할을 다룬 연구에서, 무균 마우스는 저농도 지방에 대한 선호도가 증가하고 지방으로부터의 칼로리 섭취가 증가했다 [145]. 흥미롭게도, 이러한 마우스는 미뢰에서 CD36 발현도 증가했지만, 장에서 CD36 발현은 감소했다 [145]. 섭취한 무균 마우스의 미뢰에서 CD36 발현 수준은 공복 대조군 마우스와 유사했는데, 이는 무균 마우스가 에너지 고갈 상태에 있었음을 시사한다 [16]. 이러한 마우스에서 자당 소비 증가와 유사하게, 지방에 대한 선호도 증가는 만성 에너지 결핍 상태에 대한 적응으로, 마른 체형 표현형을 유발할 수 있다. 무균 마우스는 또한 장에서 포만감 펩타이드(satiety peptides)인 CCK, GLP‑1, 그리고 PYY의 단백질 수치가 감소했으며, 이는 GPR40, 유리 지방산 수용체 3 (GPR41), 유리 지방산 수용체 2 (GPR43), 그리고 GPR120의 발현 감소와 함께 나타났다 [145]. 이러한 결과는 장내 미생물이 장의 지방산 수용체 수치 조절에 관여함을 시사한다. 순환 렙틴 및 PYY 수치도 무균 마우스에서 감소했으며 [145], 이는 이러한 동물의 마른 표현형과 일치한다. 놀랍게도 무균 마우스는 순환 그렐린 수치가 감소했는데, 이는 만성 에너지 결핍 조건에서 증가할 것으로 예상된다.
Chemosensory processes in the pancreas
췌장에서의 화학감각적 프로세스
膵臟(pancreas)의 관 시스템에 있는 Brush cells(솔세포, 刷状细胞)는 α‑ gustducin (Figure 4)을 함유하고 있으며, 이 물질은 小葉管導管 (interlobular ducts)의 말단과 주요 膵臟管(pancreatic duct) 표면에 집중되어 있다 [147]. 췌장관에 고전적 미각 수용체 분자가 존재하는 것의 기능적 중요성은 아직 알려져 있지 않다. 그러나 brush cells가 腺胞細胞 분비물(acinar secretions) 및/또는 관 상피(ductal epithelium) 자체에 존재하는 분자에 있는 화학 물질을 감지한다고 제안하는 것은 타당해 보인다. β 세포에 α‑gustducin이 존재한다는 것은 잘 알려져 있으며, 초기 연구에서는 쓴맛을 내는 denatonium benzoate를 도구로 사용하여 인슐린 분비에 대한 잠재적인 관여를 설명했다 [148]. 데나토늄 벤조에이트는 마우스 膵島(islets)에서 포도당-매개 인슐린 분비를 증가시켰지만, 그 효과는 α‑ gustducin 활성화에 기인할 수 없었다. 오히려 쓴맛을 내는 데나토늄 벤조에이트가 KATP 채널의 활성을 감소시켜 췌도 β 세포의 탈분극과 Ca2+ 유입 증가를 초래했다.
TAS1R은 β 세포에도 존재한다 [149]. 마우스 β‑세포 라인에서 인공 감미료(sucralose, acesulfame potassium 그리고 saccharin 등)에 노출되면 type II TBCs에서 TAS1R2 및 TAS1R3 이종이량체들이 활성화되고 농도 의존적으로 인슐린 분비가 자극되었다 [149]. 인공 감미료 자극은 ATP (정상적인 생리 조건에서 β 세포의 탈분극 및 인슐린 분비에 필수)를 생성하지 않는다는 점을 고려하면, TAS1R과 그 하위 신호전달 경로가 이러한 물질에 의해 활성화되어 인슐린 분비를 유도하는 것으로 추정된다. 분리된 마우스 膵島(pancreatic islets)에서 수크랄로스는 포도당 존재 하에서 인슐린 분비를 증진시키는 것으로 나타났다 [149].
果糖(Fructose)은 단맛 수용체의 강력한 활성제이며, 단맛 수용체 의존적 방식으로 포도당-매개 인슐린 분비를 증가시킨다 [150]; 그러나 이 효과는 TAS1R2 또는 TAS1R3이 결핍된 마우스에서 분리한 膵島에서는 나타나지 않았다. 과당(체중 1kg당 1g)을 정맥 주사하면 야생형 마우스에서 순환 인슐린 수치가 일시적으로 증가했지만 TAS1R2가 결핍된 마우스에서는 그렇지 않았다 [150]. 사람의 경우, 과당을 정맥 주사하면 공복 상태에서 인슐린 분비에 거의 또는 전혀 영향을 미치지 않았지만, 포도당을 정맥 주사하여 혈장 포도당 수치를 먼저 증가시켰을 때 인슐린 분비가 상당히 증가했다 [151]. 분리된 사람의 췌도에서 과당(3.0mM)은 고농도의 포도당(11.0mM)에서 포도당-유도 인슐린 분비를 증가시켰지만, 포도당 농도가 5.5mM일 때는 그렇지 않았다; 이러한 증강은 TAS1R3 수용체 알로스테릭 억제제(allosteric inhibitor)인 락티솔(lactisole)에 의해 차단되었다 [150].
이 데이터에 대한 단서는 果糖이나 蔗糖이 풍부한 음료(일반적으로 농도가 500μM 미만)를 섭취한 후에도 생체 내 과당 농도가 이 연구에서 사용된 수준에 도달하지 않는다는 것이다 [152]. 특히, 이러한 효과는 TAS1R3를 유사하게 활성화하는 사카린에서도 관찰되었다 [153]. 과당에 의한 미각 분자의 활성화를 통한 인슐린 분비 증강은 PLCβ2에 의존하는 것으로 보인다. PLCβ2의 활성이 길항제에 의해 억제되었을 때 마우스 β 세포는 과당에 반응하지 않았기 때문이다 [150].
또한, 새로운 연구 결과에 따르면 TAS1R이 기저(비-포도당 자극) 인슐린 분비에 관여하는 것으로 나타났다 [154].
TAS1R2가 결핍된 마우스의 膵島는 공복 혈당 수치에서 인슐린을 과분비하며, 이러한 마우스는 5시간 공복 후 야생형 마우스보다 혈당 수치는 낮았지만 순환 인슐린 수치는 더 높았다. 그러나 膵島의 TAS1R2 수치는 TAS1R3 수치보다 훨씬 낮았으며, TAS1R2의 녹다운은 수크랄로스 반응에 영향을 미치지 않는 것으로 나타났다 [155]. 이러한 불일치를 설명하는 한 가지 가능성은 β 세포에서 TAS1R3가 동종이량체(homodimer)로 기능할 수 있다는 것이다 (Figure 4). 사용 가능한 데이터에 대한 논란이 있기 때문에 β 세포에서 미각 수용체의 기능을 완전히 이해하기 위해서는 아직 많은 연구가 필요하다.
Neuronal control of taste
미각에 대한 신경의 컨트롤
味覺(Taste sensation)은 腦에서 양쪽으로 표현된다 [156].
Cranial nerves (腦神經) VII (facial, 顔面神經) , IX (glossopharyngeal, 舌咽神經) 및 X(vagus, 迷走神經)는 여러 중계소(relay stations)를 통해 미각 정보를 전달하며, 이 중계소는 궁극적으로 primary taste cortex (일차 미각 피질)(瓣蓋皮質에 의해 덮여 있는 島(insula)에 위치)에 연결된다. 뇌신경의 신경 섬유는 髓質(medulla)의 고립로(solitary tract, NTS)의 동측 핵(ipsilateral nucleus)으로 들어간다. 설치류에서 NTS 遠心性 纖維(efferent fibres)는 視床(thalamus)의 뉴런과 시냅스를 형성하는 腦橋(pons)의 팔곁핵(parabrachial nucleus, PBN)의 미각 중추로 미각 정보를 전달한다 (Figure 4). 靈長類(primates)에서는, NTS 섬유는 PBN을 우회하여 시상 뉴런(thalamic neurons)과 직접 시냅스를 형성한다. 설치류와 영장류 모두에서 시상 구심성 신경(thalamic afferents)은 섬엽(insula)으로 투사된다. 이어서 미각 피질(taste cortex)은 扁桃体(amygdala)로, 그리고 外側視床下部(lateral hypothalamus)와 側座核(nucleus accumbens)으로 투사를 보내는데, 여기서 쾌락적 보상 자극에 반응하여 도파민이 분비된다 [157].
PBN에서 또는 PBN 바로 위에서, 혀로부터의 맛 지각을 운반하는 상행 신경섬유(ascending nerve fibres)의 3분의 1이 교차하고, 시상 미각 영역(thalamic taste area)으로 양측으로 상행하여 뇌에서 양측 미각 표현이 가능하게 한다 [157]. NTS는 미각 정보를 전달하는 3개의 뇌신경 모두에서 나오는 미각 섬유, 장에서 나오는 자율신경 원심성 및 구심성 섬유 (autonomic efferent and afferent fibres), 그리고 뇌신경 V(삼차신경, trigeminal nerve)를 통해 얼굴, 입, 혀에서 나오는 체성 감각 구심성 섬유(somatosensory afferent fibres)가 모이는 중요한 부위이다 [158]. 또한, NTS로부터의 국소 투사는 입 안의 타액 분비 속도를 조절하는데, 이는 음식 씹기와 감상에 필요하다 [156]. 입안에 신맛이 나는 물질이 있을 경우 타액 분비가 활발하게 이루어진다 [159]. 설인신경(glossopharyngeal nerve)의 지배를 받는 Von Ebner’s glands (폰 에브너腺)은 리파아제(lipases)를 함유한 漿液 物質(serous material)을 분비하여 씹은 음식(masticated food)에서 유리지방산(GPR120 및 CD36의 리간드)이 방출되도록 한다 [160].
TAS2R5 또는 TAS1R3 프로모터에 의해 구동되는 형광 표지된 WGA를 발현하는 형질전환 마우스를 사용한 실험에서 쓴맛에 반응하는 신경 세포가 꼬리 영역(caudal regions)에서 발견되었고, TAS1R3을 전달하는 신경 세포가 NTS, PBN, 視床(thalamus) 및 미각 피질(gustatory cortex)의 전방 영역(rostral regions)에서 확인되었다 [56]. TAS2R5-WGA 형질전환 마우스만을 사용한 기능 테스트에서 형광 WGA로 표지된 뇌 영역에서 cycloheximide (쓴맛을 내는 물질)를 혀에 적용했을 때 즉각적 초기 반응 유전자인 Zif268 (조건화된 맛 혐오 학습에 관련된 생성물을 인코딩함)[161]의 발현이 증가했다 [162]. 이러한 데이터를 종합해 보면 뇌의 특정 영역이 개인의 미각에 관련된 정보를 전달하는 데 전념한다는 개념을 뒷받침하는 것으로 보인다. 그러나 비형광 WGA를 사용한 연구에서는 NTS를 넘어 쓴맛 미각 물질에 반응하는 신경 세포의 확실한 표지를 찾지 못했는데, 이는 방법론의 민감성이 부족했기 때문일 수 있다 [57,54,163]. 정교한 생체 내 2광자 칼슘 이미징(two-photon-calcium imaging)과 순행성 추적자(antegrade tracer)를 사용하여 식별된 맛 반응 세포에서부터 섬피질(insula)로의 투사를 표지하는 포괄적인 연구에서 단맛, 쓴맛, 우마미, 짠맛에 대한 미각 지각(신맛은 아님)은 섬피질(insula)에서 뚜렷한 지형적 구분(discrete topographic segregation)을 보인다는 것이 입증되었다 [164]. 일부 신경과학자들은 미각 감각 뉴런이 중추 신경계에서 광범위하게 조정된다(tuned)고 생각하기 때문에, 미각 지도(gustatory map)의 완전성에 대한 의문이 남아 있다 [165,166]. 이 데이터는 이후 혀의 TBCs로부터 섬엽(insula)의 전용 영역으로 미각 특정 정보를 전달하는 구심 신경 경로를 설득력 있게 정의하는 자세한 후속 연구[167]에서 입증되었다.
포도당 센서(Glucose sensors)는 視床下部(hypothalamus), 腦幹(brainstem), 扁桃体(amygdala), 中隔(septum), 皮質(cortex)에 존재하며, 특히 低血糖(hypoglycaemia)에 대한 역조절 반응에 관여하는 뇌 영역인 시상하부의 室傍核(paraventricular nucleus)에 풍부하게 존재한다 [168–172]. 이러한 뉴런은 포도당에 대한 논리적 반응에 따라 일반적으로 포도당 흥분성(glucose-excited) 뉴런과 포도당 억제성(glucose-inhibited) 뉴런으로 분류된다. 뉴런은 랑게르한스 섬의 β 세포와 유사한 방식으로 glucokinase/KATP-dependent pathways를 통해 포도당을 감지하는 것으로 여겨졌지만[172], 전형적인 단맛 수용체와 α‑gustducin은 전뇌(forebrain)의 여러 영역(視床下部(hypothalamus), 海馬(hippocampus), 하베눌라(habenula, 고삐), 脈絡叢(choroid plexus, 脈絡膜網), 그리고 皮質(cortex) 포함)에서 발현된다 [173,174]. 더욱이 뇌에서 단맛 수용체의 발현 수준은 동물의 영양 상태에 의해 조절된다. 視床下部에서 영양 결핍은 단맛 수용체의 발현 증가를 초래하며, 이 효과는 저-포도당 조건에서 배양한 마우스 시상하부 세포 라인에서 재현되었다. 흥미롭게도, 이 효과는 세포에 수크랄로스(대사되지 않는 일반적으로 사용되는 감미료)를 첨가하자 역전되었다 [175]. 이러한 연구 결과를 종합해 보면, 포도당 대사와 그에 따른 ATP 생성이 시상하부 신경 세포에서 이러한 수용체의 발현을 조절하는 데 필요하지 않다는 것을 알 수 있다.
비교해보면, 비만 마우스(과영양(overnutrition)과 혈당과다(hyperglycaemia)의 모델이 될 수 있음)에서는 시상하부의 단맛 수용체 발현 수치가 감소했다 [173]. 게다가 인지 기능에 중요한 海馬 뉴런은 α‑gustducin, TAS1R2, 그리고및 TAS1R3을 지속적으로 공동 발현한다 [173,174]. 마우스에게 식수에 감미료인 아세설팜칼륨(acesulfame potassium)을 투여한 연구에서는, 해마에서 TAS1R3 발현 수치가 감소한 것으로 나타났는데, 이 현상은 전형적인 포도당 수송체 수치의 변화와 인지 장애의 감소와 병행되었다 [174]. 또한 마우스에서 TAS1R3을 제거(ablation)하면 영양소-유도 mTORC1 활성화(nutrient- induced activation of mTORC1)가 감소했는데, 이는 TAS1R이 영양소 상태를 측정하는 세포 신호 전달 메커니즘의 상류에서 기능한다는 것을 시사한다 [176]. 이러한 결과를 바탕으로 우리는 뇌의 미각 수용체가 영양 상태, 대사 지원, 세포 회전율(degree of cell turnover) 등을 장기적으로 감지하는 시스템의 일부라고 제안한다.
糖의 테이스팅(tasting)과 소화(digesting)(postabsorption)가 이중 보상(dual reward)을 제공한다는 것은 잘 알려져 있다. 보상 시스템은 시냅스 후 뇌간 경로(postsynaptic brainstem pathway)를 통해 맛이 좋은 음식을 맛볼 때 처음 활성화되고 흡수 후에 재활성화된다 [177]. 적어도 마우스들에서는 칼로리가 높은 음식의 흡수가 맛 지각(taste perception)과는 별개로 보상 시스템에 직접적인 영향을 미치는 것으로 보인다[177–179]. 예를 들어, TBC에서 단맛, 우마미, 쓴맛 테이스팅 신호를 전달할 수 없는 Trpm5-/- 마우스들은 蔗糖 음료에 대한 강한 선호도를 보인다 [177]. 더 나아가, 자당을 섭취하면 야생형 마우스들과 Trpm5-/- 마우스들 모두에서 側座核 (nucleus accumbens)과 腹部 腺條體 (ventral striatum)에서 도파민(dopamine)이 분비되었다 [180]. 세포의 포도당 이용을 방해하는 분자인 2-deoxyglucose(데옥시글루코스를)를 투여하여 포도당 산화를 방해하면 칼로리가 함유된 음식을 찾는 마우스에게 인공감미료 섭취를 억제하는 효과가 있었는데, 이는 쾌락적 보상을 위한 포도당 산화의 필요성을 보여준다 [181]. 반대로, 소화 후 효과에 반응하여 발생하는 도파민 방출을 억제하면 포도당 결핍 조건에서도 포도당 섭취가 감소했다 [181]. 도파민 뉴런의 활성화는 수크랄로스의 맛과 상승 작용을 하며 수크로스만 섭취했을 때보다 더 높은 가치의 총 자극을 생성한다 [182]. 포도당 섭취 전에 렙틴(leptin)을 투여하면 도파민 활성화가 감소하고 설탕 섭취가 감소했으며, 따라서 렙틴은 음식에 대한 쾌락적 반응의 중요한 조절자(regulator)이며 영양소의 보상 가치 조절자이기도 한 것으로 보인다 [182].
Taste perception and obesity
맛 지각과 비만
저렴하고 맛이 좋은 고칼로리 가공식품의 증가로 비만 유병률이 증가했다. 비만이 진행됨에 따라 맛 지각에도 변화가 나타나는 것으로 여겨진다. 마우스 연구에 따르면, 고지방 식단을 섭취한 동물은 일반 사료를 섭취한 마우스보다 미뢰에 단맛 TBCs가 더 적었고, 단맛 물질과 LCFAs에 대한 반응도 감소했지만, 우마미 물질에는 변화가 없었다 [183]. 생리학적으로 고지방 식단을 섭취한 마우스는 마른 마우스보다 유사한 반응을 유발하기 위해 단맛 물질과 지방 맛 물질이 더 많이 필요했다. 이러한 맛 지각의 조절(modulation)은 고지방 식단이 미각 수용체 세포의 분포와 수에 변화를 일으킬 수 있음을 시사한다. 소아에서 지방 맛에 대한 선호도와 체질량 지수(BMI) 사이에는 양의 상관관계가 보고되었다 [184]. CD36과 달리 GPR120의 발현은 지방이나 혼합 식품의 경구 섭취에 대한 반응으로 하향 조절되지 않는다 [16]. 이는 이 수용체가 섭취한 음식의 양(칼로리 상당량)을 맛 지각과 그리고 장, 간, 지방 조직 및 뇌의 반응과 통합하는 생리학적 '하드 드라이브'일 수 있음을 시사한다.
지난 천년 이전에 희소했던 설탕이 첨가된 식품이 도입되기 전에는, GPR120이 가공되지 않은 식품에서 얻어야 하는 신체의 에너지 요구를 충족하는 매우 중요한 통합자였을 것이다. 이 기능은 현대 식단에 칼로리가 높은 가공 식품이 포함되면서 이제 덜 중요해졌다. GPR120이 결핍된 마우스에게 고지방 식단을 제공하면 비만과 포도당 불내증(glucose intolerance)이 발생했다 [185]. 동물 모델에서의 결과와 일치하게, GPR120에 non-synonymous mutation를 가지고 있어 신호전달 활동에 결함이 있는 사람은 비만 유병률이 증가했다 [185]. 중요한 것은 건강한 개인의 경우 지방 형태로 단독으로 섭취하는 LCFAs(단맛이나 소금을 첨가하지 않음)는 식욕을 돋우지 않기 때문에 과식에 도움이 되지 않는다는 것이다.
Surgical interventions affecting taste and food intake
맛과 음식 섭취에 영향미치는 수술적 개입
Weight-loss surgery
대부분의 위와 근위 장(proximal intestine)이 음식과의 접촉으로부터 분리되는 루-앙-와이 胃 우회술(RYGB)은 비만 치료에 널리 사용되는 절차이다. RYGB 수술 후 환자는 체중이 60~70% 감소할 수 있으며, 이는 최대 15년 동안 유지될 수 있다 [186,187]. 또한, 제2형 당뇨병과 같은 일부 비만 관련 대사 장애는 수술 후 다양한 기간 내에 60~80%의 寛解率(remission rate, 緩解率)로 해결될 수 있다 [186,187].
쥐를 대상으로 한 연구에서는 RYGB 후 체중이 장기적으로 상당히 감소했으며 [188~191], 이는 고농도의 수크로스에 대한 선호도 감소와 관련이 있었으며, 수술군과 대조군에서 저농도의 수크로스에 대한 선호도에는 차이가 없었다 [189]. 또 다른 연구에서는 RYGB 후 쥐의 수크로스 선호도 점수가 가짜 수술을 받은 쥐(sham-operated rats)보다 높았다는 것을 발견했다. 그러나 구연산(신맛), 염화나트륨(저염), 키닌(쓴맛) 선호도에는 차이가 관찰되지 않았다 [190].
RYBG 수술의 설치류 모델에서 식욕에 영향을 줄 수 있는 순환 GLP‑1 및 PYY 수치의 증가가 일관되게 입증되었다 [190–193]. 그러나 GLP‑1 수치의 상당한 증가에도 불구하고, 기능적 GLP‑1 결핍의 다양한 동물 모델(예: GLP‑1 수용체 길항제로 치료한 마우스, 1-수용체 결핍 마우스 및 α‑거스트듀신 결핍 마우스)도 위 우회 수술 후 가성 수술을 받은 동물에 비해 체중이 감소하고 포도당 항상성이 향상되었다 [192,193]. 설치류 실험 결과는 위 우회 수술로 치료한 결과 고칼로리 음식에 대한 선호도가 감소한 인간 실험 결과와 일치한다 [194,195].
RYGB 수술(82명)과 복강경 위 밴드술 (laparoscopic gastric banding, LGB, 28명)이 수술 후 음식의 즐거움에 미치는 영향을 조사한 초기 연구에서, 많은 개인들이 맛 강도 감소를 보고했다. 또한, RYGB 치료를 받은 사람의 67%와 LGB 수술을 받은 사람의 68%가 쓴맛과 신맛이 나는 음식과 같은 특정 음식에 대한 혐오감을 느꼈고, 단맛에 대한 미각도 증가했다. 게다가, 대부분의 연구 참여자들은 미각 변화가 체중 감량에 도움이 된다고 느꼈다 [196]. 대사 수술 후 식품의 보상 특성 변화를 평가한 연구에서는 RYGB 수술을 받은 환자에서 대조군에 비해 초콜릿에 대한 선호도가 감소하고 채소에 대한 선호도가 증가한 것으로 나타났다 [194]. 또한, RYGB 수술 후 체중이 약 20% 감소한 여성은 단맛에 대한 기호도가 즐거운 것에서 불쾌한 것으로 빠르게 바뀌었다고 보고했으며, 이로 인해 단 음식 섭취가 감소했다 [195]. 대사 수술이 미뢰 형태 또는 특정 TBCs 집단이나 호르몬에 영향을 미치는지는 아직 연구되지 않았다.
Vagotomy 迷走神經切斷術
뇌에서 미주신경 입력의 주요 투사 부위는 미각신경(gustatory nerves)의 투사 부위이기도 하다. 설치류 모델에서 미주신경 절단술(즉, 복부에서 미주신경을 절제하는 수술)에 대한 연구에 따르면, 미주신경은 장이 미각 음식에 대한 혐오감과 끌림을 조절하는 데 필수적이다 [121,178,179,197]. 쥐에서 횡격막하 미주신경 절단술(subdiaphragmatic vagotomy) 후, 자당과 사카린 섭취는 감소했지만, 물 섭취량은 변화가 없었다 [198]. 흥미롭게도, 미주신경 절단술 후 쥐의 음식 섭취량과 체중이 감소했다 [199]. 쥐에서 양측 몸통 미주신경 절단술을 완전히 시행한 결과, 수술 후 4주 동안 음식 섭취량이 20% 감소했으며, 혈청 렙틴 수치와 총 체지방 감소가 동반되었다. 그러나 양측 장내신경 절단술을 시행한 쥐에서는 체중 감소가 나타나지 않았다 [200]. 인간의 경우, 체간 미주신경절제술(truncal vagotomy) 후 체중 변화에 대한 데이터는 상충되는데, 이는 미주신경 절제술의 불완전한 진행 및/또는 미주신경 섬유의 부분적인 재성장의 결과일 수 있다. 미주신경절제술 쥐 모델 연구에서도 장 유래 엔도카나비노이드(gut-derived endocannabinoids) 수치가 감소한 것으로 보고되었으며 [141,142], 미주신경절제술이 CCK 및 GLP-1과 같은 음식 섭취에 영향을 미치는 다른 體液性 因子(humoral factors)의 분비에 영향을 미칠 가능성이 있다. 특히, 미주신경절제술이 장이나 미뢰의 단맛, 쓴맛 또는 지방 수용체 발현에 미치는 영향은 보고된 바가 없다.
Conclusions
쓴맛, 고염분, 신맛은 잠재적으로 치명적인 화합물 섭취를 피하고 방어하는 역할을 한다. 단맛과 감칠맛은 식품의 영양소 함량을 반영하지만, 비영양 인공감미료의 등장으로 이러한 관계가 항상 일치하는 것은 아니다. 맛 지각을 음식의 섭취 후 특성('기분 좋은' 측면)과 분리해도 적어도 동물 연구에서는 체중 감소로 이어지지 않으며, 이는 음식의 쾌락적 가치의 중요성을 시사한다. 음식에 단맛, 약간의 소금, 그리고 약간의 신맛을 더하면 기호성과 씹는 맛이 증가하여 쾌락적 가치가 증가한다. 저렴한 고-과당 옥수수 시럽의 보급으로 설탕 사용이 증가했고, 1회 제공량 증가와 음식 접근성의 용이성과 맞물려 시간이 지남에 따라 음식의 쾌락적 가치가 포만 신호를 무시하게 될 수 있다. 음식을 준비하고 적절한 칼로리를 섭취하는 데 필요한 작업량이 감소함에 따라 현대 사회의 음식 섭취량도 영향을 받았으며, 이로 인해 비만이 만연하는 속도가 더욱 빨라졌다.
비만 자체가 맛과 맛 지각에 영향을 미칠 가능성도 있다. 그러나 인간의 맛 지각에 관한 자료는 상충되는 것으로 보인다. 비만이었던 사람들이 체중 감량 수술이 아닌 다이어트로 체중을 감량한 경우, 마른 사람들에 비해 단 음식과 고지방 음식에 대한 욕구가 더 높다는 증거가 있는데, 이는 중추 신경계의 에너지 부족으로 인해 칼로리 제한 식단을 유지하기 어렵기 때문일 수 있다 [201]. 맛 지각의 성별 차이도 보고되었지만 [202], 이는 인간을 대상으로 한 복잡한 미각 연구에서 항상 고려된 것은 아니다. 요약하자면, 과도한 에너지 섭취는 맛, 쾌락 반응, 그리고 뇌와 말초로의 섭취 후 피드백 루프 간의 상호작용을 수반하는 다면적인 대사 상태이다. 따라서 과도한 에너지 섭취와 관련된 부작용을 해결하기 위해서는 피드백 루프에 개입하는 새로운 접근법이 시급히 필요하다.
References




'Coffee Physiology' 카테고리의 다른 글
| 혀의 生理學 – 테이스트 信號傳達 [1] (3) | 2025.06.30 |
|---|---|
| 味蕾에서의 테이스트 전달과 채널 시냅스 (29) | 2025.06.19 |
| 味蕾 : 細胞, 信號 그리고 시냅스 (20) | 2025.06.15 |
| 味覺 細胞 生物學 (15) | 2025.06.07 |
| 에스프레소 커피 知覺의 生理學 (6) | 2025.06.05 |



